Эволюция геосфер и биосферы¶
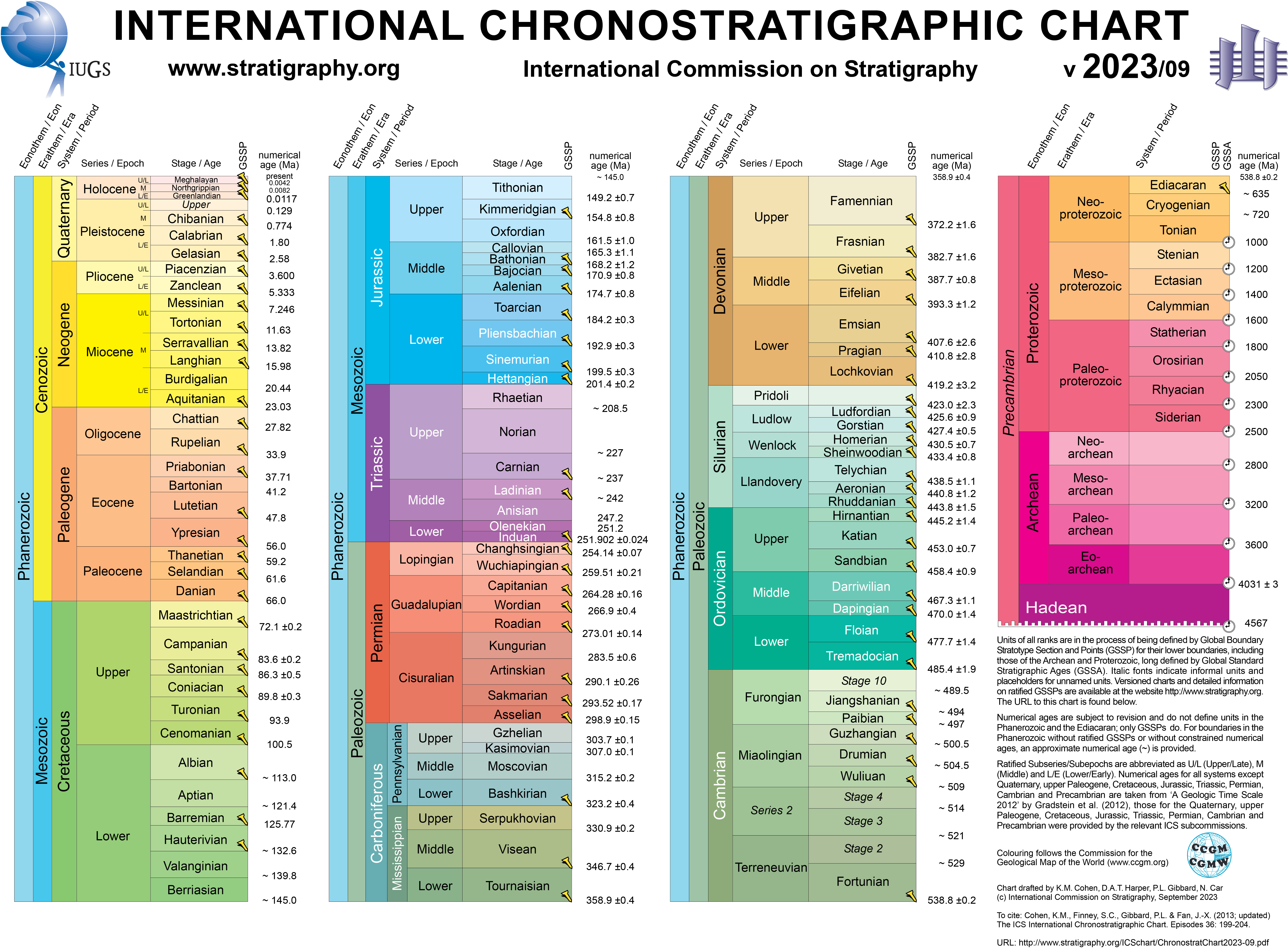
Хроностратиграфическая (геохронологическая) шкала¶
Хроностратиграфическая шкала (геохронологическая шкала) — геологическая временнáя шкала истории Земли, применяемая в геологии и палеонтологии, своеобразный календарь для промежутков времени в сотни тысяч и миллионы лет.
Согласно современным общепринятым представлениям возраст Земли оценивается в 4,5–4,6 млрд лет. На поверхности Земли не обнаружены горные породы или минералы, которые могли бы быть свидетелями образования планеты.
Максимальный возраст Земли ограничивается возрастом самых ранних твёрдых образований в Солнечной системе — тугоплавких включений, богатых кальцием и алюминием из углистых хондритов. Возраст таких включений из метеорита Альенде по результатам современных исследований уран-свинцовым методом составляет 4,56 млн лет. Время формирования Земли как планеты может быть позже этой даты на миллионы и даже многие десятки миллионов лет.
Хондриты — одна из подгрупп метеоритов, которые составляют более 92 % упавших на Землю метеоритов.

Последующее время в истории Земли, после предполагаемой даты её образования, было разделено на временные интервалы разной длительности. Их границы проведены по важнейшим тогда происходящим событиям.
Во второй половине XIX века на II–VIII сессиях Международного геологического конгресса в 1881–1900 годах были приняты иерархия и номенклатура большинства современных геохронологических подразделений. В последующем Международная геохронологическая шкала постоянно уточнялась.

Конкретные названия периодам давали по разным признакам. Чаще всего использовали географические названия. Так, название кембрийского периода происходит от лат. Cambria — названия Уэльса, когда он был в составе Римской империи, девонского — от графства Девоншир в Англии, пермского — от г. Перми, юрского — от гор Юра в Европе. В честь древних племён названы вендский, ордовикский и силурийский периоды. Реже использовались названия, связанные с составом пород. Каменноугольный период назван из-за большого количества угольных пластов, а меловой — из-за широкого распространения мела.
Геохронологическую шкалу создавали для определения относительного геологического возраста пород. Абсолютный возраст, измеряемый в годах, имеет для геологов второстепенное значение.
Время существования Земли разделено на два главных интервала: фанерозой и докембрий (криптозой) по появлению в осадочных породах ископаемых остатков.
Докембрий — время скрытой жизни, в нём существовали только «мягкотелые» организмы, не оставляющие следов в осадочных породах.
Фанерозой начался с появлением на границе эдиакария (венд) и кембрия множества видов моллюсков и других организмов, позволяющих палеонтологам расчленять толщи по находкам ископаемой флоры и фауны.
Развитие биосферы¶
Периодизация развития биосферы¶
По В.И. Вернадскому, крупные этапы в эволюции биосферы были связаны, как правило, с появлением новых групп организмов, выполняющих специфические геохимические и энергетические функции в экосистемах. Он отмечал четыре крупных этапа в истории биосферы:
- появление первичных автотрофов,
- возникновение животных с кальциевым скелетом,
- формирование лесных биогеоценозов,
- формирование ноосферы.
В.В. Ковальский выделил в развитии живой природы:
- протобиосферу,
- биосферу,
- ноосферу.
Он рассмотрел особенности биологического круговорота на каждом из них. Однако первый и последний этапы развития биосферы выходят за рамки биоистории.
Д.И. Сапожников предложил при выделении основных этапов в эволюции живого исходить из эволюции типов обмена веществ:
- первичные гетеротрофы: эволюция типов питания на первом этапе состояла в постепенном преобразовании гетеротрофов, способных усваивать соединения, только очень богатые энергией, в гетеротрофы, способные использовать продукты ассимиляции первичных форм;
- гетеротрофные усвоители CO2: на втором этапе возникла авто-гетеротрофность, то есть автотрофность в отношении аминокислот и витаминов при способности усваивать аммиачный азот;
- хеморедукторы: с возникновением хеморедукции усвоение углекислого газа осуществлялось за счёт энергии неорганического вещества в анаэробных условиях;
- фоторедукторы: на четвёртом этапе появилась способность к фоторедукции (использование световой энергии для различных окислительно-восстановительных реакций);
- фотосинтетики: становление фотосинтеза — повышение окислительно-восстановительного потенциала, появление и совершенствование железосодержащих ферментных систем, включение в обмен свободного молекулярного кислорода.
Однако в концепции этого автора нет связи эволюции типов обмена веществ с коренными переменами в биосфере.
М.А. Голубец выделяет следующие наиболее крупные переломные этапы в истории:
- этап гетеротрофной биосферы в условиях восстановительной обстановки,
- этап существования организмов в условиях окислительной обстановки,
- этап антропогенного воздействия на процессы биосферы.
Рассматривая историю кислорода в атмосфере, Г.А. Заварзин назвал следующие этапы доминирования в биосфере геохимических процессов от настоящего времени к прошлому:
- циклический механизм, обусловленный высшей наземной растительностью и грибами, а также животными, ведущий к формированию гумуса за счёт лигноцеллюлозы с нижней границей около 0,4 млрд лет назад;
- циклический механизм, обусловленный эукариотными водорослями и беспозвоночными океана примерно 1,5–2 млрд лет назад;
- цианобактериальные сообщества и их анаэробное разложение, формирование строматолитов, окисление резервуаров восстановленных веществ — до 3,5 млрд лет назад;
- фотохимические реакции в примитивной атмосфере у поверхности Земли.
Вышеизложенное показывает, что для периодизации истории биосферы не выработаны единые критерии.
Поярков Б.В. выделяет шесть этап в эволюции биосферы. Первым он указывает добиосферный этап, что обосновано, так как именно условия, сформировавшиеся на добиосферном этапе, позволили возникнуть жизни.
Однако предложенное им подразделение на этапы имеет ряд недостатков:
- упускаются из вида множество ключевых событий внутри каждого из этапов;
- поверхностно рассматриваются происходящие события;
- нет связи этапов с тектоникой плит;
- отсутствует дифференцированное рассмотрение событий (по геосферам, климату и другому).
Далее эти недостатки нужно учитывать.
Добиосферный этап развития Земли (до появления жизни)¶
Геологическое время¶
Добиосферный этап развития Земли включает в себя катархейский эон, предшествовавший архею.
Катархей, архей и протерозой являются эонами, которые согласно хроностратиграфической шкале объединяют также в эон — докембрий. Ввиду отсутствия каких-либо более подробных геологических сведений, данный эон не подразделяют на более мелкие подразделения.
Начался данный период с формирования Земли примерно 4600 млн лет назад и продолжался до 4000 млн лет назад (всего 600 млн лет), примерно 15–25 % все историипланеты.
В этот же период сформировалась Луна — естественный спутник Земли, вероятнее всего после столкновения с планетой Тейя размером с Марс около 4360 млн лет назад. В результате такого крупного столкновения, во-первых, изменился наклон земной оси (до 23°), во-вторых, произошло массовое плавление мантии с образованием магматического океана с глубиной до 700 км.
После столкновения большая часть горных пород планеты должна была испариться, и из них вокруг планеты появилась бы атмосфера. Испарившиеся горные породы конденсировались, в результате чего оставались горячие летучие вещества, которые, вероятно, образовали тяжёлую углекислую атмосферу с водородом и водяным паром. Жидкая вода океанов существовала несмотря на поверхностную температуру 230 °C из-за давления тяжёлой атмосферы. Охлаждение продолжалось, в атмосфере значительно снизилось количество углекислого газа из-за субдукции и растворения в воде океанов.
К концу этого периода наряду с затвердеванием земной коры вследствие охлаждения планеты, возникает атмосфера и океан
Литосфера¶
К концу катархея сформировался лишь базальтовый слой земной коры за счёт потоков лав основного состава. Литогенез был в основном представлен его вулканогенно-осадочным типом. Это были, прежде всего, лавы и массы рыхлого пеплового материала.


Гидросфера¶
Химический состав «первозданного» океана, вероятно, отличался от современного. Воды океана были очень кислыми и являлись по существу крепким раствором соляной кислоты (HCl), фтороводородной кислоты (HF), борной кислоты (H3BO3), диоксида кремния (SiO2) с pH, близким к 1–2. В этой воде были растворены также и некоторые газы — сероводород (H2S), метан (CH4) и другие углеводороды, а также CO2, но сульфатов ещё не было или они существовали только в следах. Свободный кислород, который выделялся при подводных излияниях базальтов, тут же расходовался для окисления сероводорода (H2S) в оксид серы (SO2). Однако важен сам факт присутствия, пусть и весьма кратковременного, геохимической энергии свободного кислорода, выделявшегося при подводных излияниях базальтов.
К концу начального этапа в результате интенсивно протекавших процессов нейтрализации кислые — за счёт сильных кислот — океанские воды превратились в воды хлоридного типа, которые отличались от современных вод этого типа обилием хлоридов алюминия (Al), железа (Fe), марганца (Mn), полным отсутствием растворенных карбонатов и ничтожным — сульфатов. Газовая фаза этих вод была представлена главным образом C2O отчасти N2, H2 при полном отсутствии свободного кислорода. Иначе говоря, хотя процесс нейтрализации сильных кислот и сопровождался несомненным подъёмом pH, однако, и по завершении его воды оставались ещё значительно кислыми; только носителями кислотности стали уже не сильные кислоты, а слабые — прежде всего, конечно, угольная кислота (Н2СО3), потом борная кислота (H3BO3) и др.
Суша ещё была очень невелика (всего около 2 %), но внешний круговорот воды уже начал осуществляться, благодаря которому поддерживались термодинамические условия, приемлемые для существования живого вещества
Атмосфера¶
Первичная атмосфера состояла из газообразных продуктов, выделяющихся при вулканических извержениях. В составе газов находились пары воды, метан, углекислота, аммиак, азот, водород с примесью инертных газов (аргон (Аг), криптон (Кг), ксенон (Хе), гелий (Не)) и так называемые кислые дымы, выделяемые вулканами (фтороводород (HF), хлороводород (HCl), борная кислота (H3BO3), сероводород (H2S) и другие). В этой атмосфере древнего типа преобладали аммиак (NH3), метан (CH4), углекислый газ (CO2). Свободный кислород в атмосфере отсутствовал.
Происходила медленная трансформация газовой смеси в направлении накопления в ней главного компонента современной атмосферы — азота. Одновременно накапливались радиогенные инертные газы аргон (Аг), криптон (Кг), ксенон (Хе), неон (Ne), гелий (Не).
В итоге в газовой оболочке нашей планеты появились первые защитные уровни, которые смягчали воздействие Космоса на поверхность Земли.
Происхождение жизни¶
Касательно эволюции непосредственно биосферы важное значение имеет вопрос о происхождении жизни. Жизнь появилась на планете Земля около 4,1–3,8 млрд лет назад.
Ответ на вопрос о происхождении жизни человечество искало еще с глубокой древности. Первые попытки ответить на этот вопросы были связаны с представлениями, что жизнь может самозарождаться из неживого вещества.
Согласно теории самозарождения жизни некоторые частицы содержат некое активное начало, которое при подходящих условиях может создать живой организм. Распространена эта теория была в Древнем Китае, Вавилоне и Древнем Египте. Аристотель, также был приверженцем данных воззрений и не сомневался в самозарождении лягушек, мышей и других организмов.
Тосканский врач Франческо Реди в 1668 году подверг сомнению теорию спонтанного зарождения и своими экспериментами с накрыванием горшочков с мясом марлей, подтвердил, что при отсутствии контакта мух с мясом личинки в мясе не зарождаются.

Несмотря на эти эксперименты теория самозарождения продолжала оставаться основной.
Далее данной проблемой занялся французский химик Луи Пастер в 1860 году.
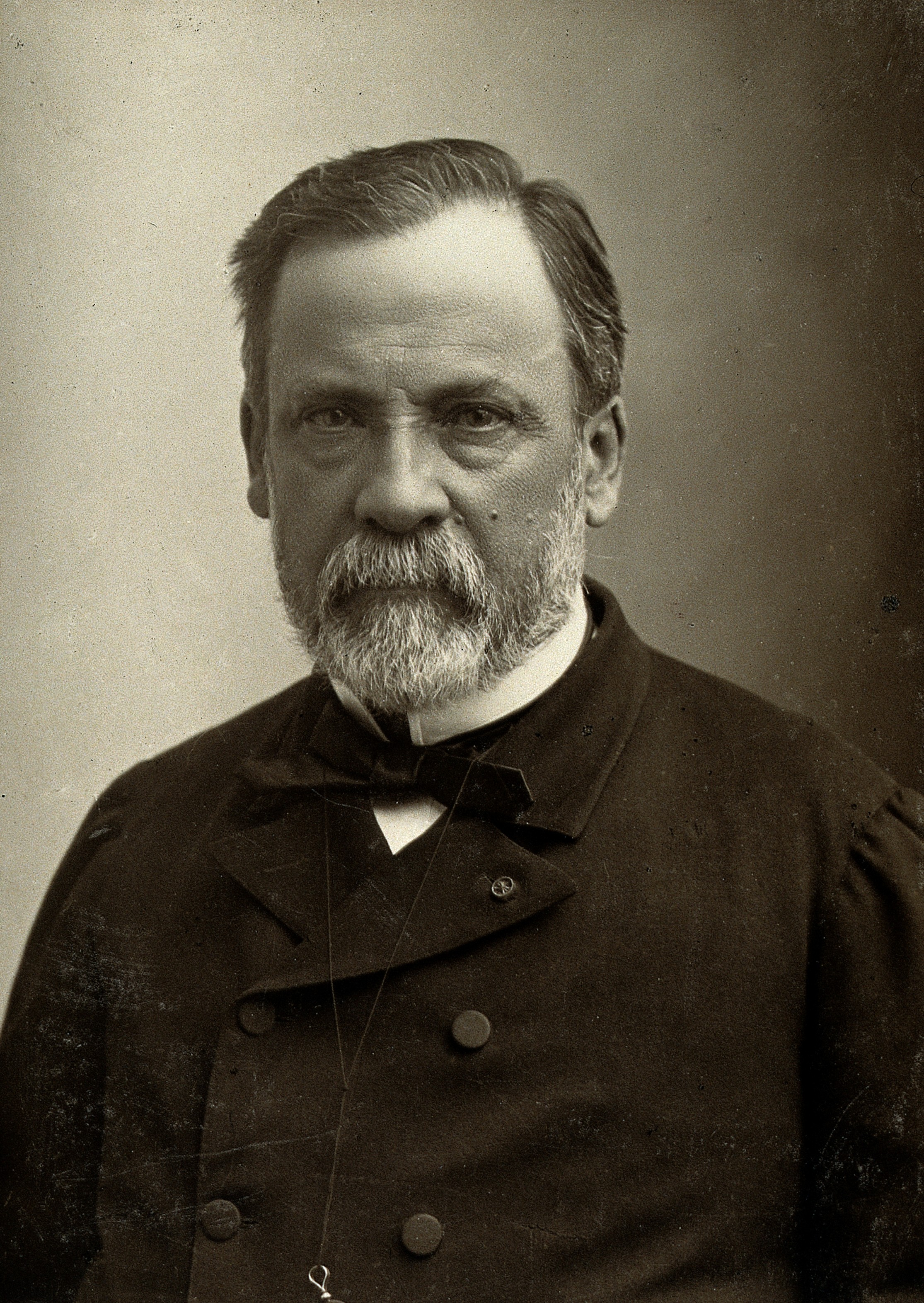
Цель опровергнуть теорию самозарождения перед собой Пастер не ставил, так как он интересовался данным вопросом для понимания связи зарождения микроорганизмов и борьбы с инфекционными заболеваниями. Т. е. стоял вопрос — есть ли смысл бороться с микроорганизмами, если они сами снова могут самозародиться. Известный эксперимент с S-образной трубкой у колбы, которая, несмотря на непрепятствие проникновения воздуха в колбу к стерильному раствору, не позволяла проникать микроорганизмам, которые оседали на её стенках.
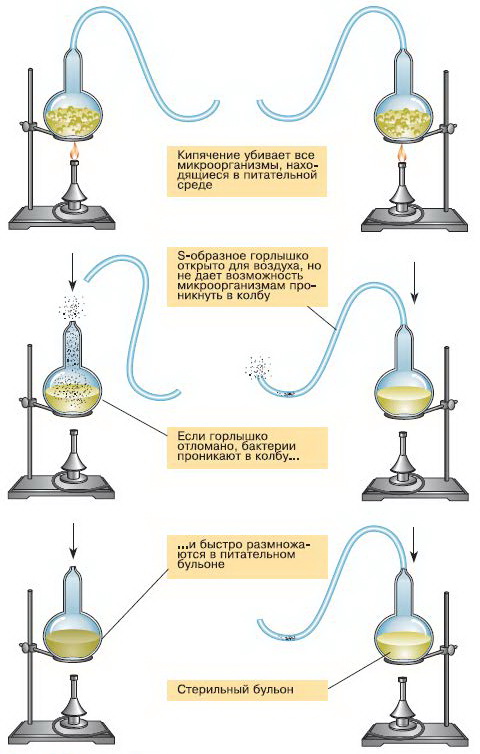
Вывод по результатам экспериментов Луи Пастера в том, что жизненной силы не существуют и в настоящее время микроорганизмы не самозарождаются из неживого субстрата.
Опыты Ф. Реди и Л. Пастера и другие сходные опыты убедительно показали, что в современную эпоху живые организмы происходят только от других живых организмов. Невозможность самозарождения жизни из неживого назвали принципом Реди. И в результате закономерен вопрос о происхождении первых живых организмов.
По вопросу происхождения жизни, так же как и по вопросу о сущности жизни, среди ученых нет единого мнения. Существует несколько подходов к решению вопроса о происхождения жизни, которые тесно переплетаются между собой. Классифицировать их можно следующим образом:
- по принципу, что идея и разум первичны, а материя вторична (идеалистические гипотезы) или материя первична, а идея и разум вторичны (материалистические гипотезы);
- по принципу, что жизнь существовала всегда и будет существовать вечно (гипотезы стационарного состояния) или жизнь возникает на определенном этапе развития мира;
- по принципу «живое только от живого» (гипотезы биогенеза) или возможно самозарождение живого из неживого (гипотезы абиогенеза);
- по принципу, что возникла на Земле или была занесена из космоса (гипотезы панспермии).
Креационизм
Согласно этой гипотезе жизнь была создана Творцом. Творец — Бог, Идея, Высший разум или другое.
Гипотеза стационарного состояния
Жизнь, как и сама Вселенная, существовала всегда и будет существовать вечно, так как не имеющее начала не имеет и конца. Вместе с тем существование отдельных тел и образований (звезд, планет, организмов) ограничено во времени, они возникают, рождаются и погибают. В настоящее время эта гипотеза имеет в основном историческое значение, так как общепризнанной теорией образования Вселенной является теория Большого взрыва, согласно которой Вселенная существует ограниченное время, она образовалась из одной точки около 15 млрд лет назад.
Гипотеза панспермии
Жизнь на Землю была занесена из космоса и прижилась здесь, после того как на Земле сложились благоприятные для этого условия. Это предположение высказал немецкий ученый Г. Рихтер в 1865 году, окончательно сформулировал шведский ученый С. Аррениус в 1895 году. С метеоритами и космической пылью на Землю могли попасть споры бактерий, которые в значительной степени устойчивы к радиации, вакууму, низким температурам. Решение вопроса о том, как возникла жизнь в космосе, в силу объективных трудностей его решения отодвигается на неопределенное время. Она могла быть создана Творцом, существовать всегда или возникнуть из неживого вещества.
Гипотеза абиогенеза (самозарождения живого из неживого и последующей биохимической эволюции). Теория Опарина-Холдейна В 1924 году Александр Иванович Опарин предположил, что в растворах высокомолекулярных соединений могут самопроизвольно образовываться зоны повышенной концентрации, которые относительно отделены от внешней среды и могут поддерживать обмен с ней. Он назвал их коацерватные капли, или просто коацерваты. Подобные взгляды также высказывал британский биолог Джон Холдейн в 1929 году.

Согласно этой гипотезе в процессе становления жизни можно выделить четыре этапа:
- Синтез низкомолекулярных органических соединений из газов первичной атмосферы;
- Полимеризация мономеров с образованием цепей белков и нуклеиновых кислот;
- Образование фазовообособленных систем органических веществ, отделенных от внешней среды мембранами;
- Возникновение простейших клеток, обладающих свойствами живого, в том числе репродуктивным аппаратом, осуществляющим передачу дочерним клеткам всех химических и метаболических свойств родительских клеток.
Первые три этапа относят к периоду химической эволюции, с четвертого начинается биологическая эволюция.
В 1953 году теорию проверил Стэнли Миллер в эксперименте Миллера-Юри. Он поместил смесь водорода (H2), воды (H2O), аммиака (NH3), метана (CH4), углекислого газа (CO2), монооксида углерода (угарный газ) (CO) в замкнутый сосуд и стал пропускать через неё электрические разряды (при температуре 80 °С). Оказалось, что образуются аминокислоты. Позднее в разных условиях были получены также сахара и нуклеотиды. Он сделал вывод, что эволюция может произойти при фазовообособленном состоянии из раствора (коацерватов).

Теория была обоснована, кроме одной проблемы, на которую долго закрывали глаза почти все специалисты в области происхождения жизни. Если спонтанно, путём случайных безматричных синтезов в коацервате возникали единичные удачные конструкции белковых молекул, то как они могли копироваться для распространения внутри коацервата, а тем более для передачи коацерватам-потомкам? Теория оказалась неспособной предложить решение проблемы точного воспроизведения — внутри коацервата и в поколениях — единичных, случайно появившихся эффективных белковых структур.

К XXI веку теория Опарина-Холдейна, предполагающая изначальное возникновение белков, практически уступила место более современной. Толчком к её разработке послужило открытие рибозимов («рибонуклеиновая кислота» и «энзим») — молекул РНК, обладающих ферментативной активностью и поэтому способных соединять в себе функции, которые в настоящих клетках в основном выполняют по отдельности белки и ДНК, то есть катализирование биохимических реакций и хранение наследственной информации. Таким образом, предполагается, что первые живые существа были РНК-организмами без белков и ДНК, а прообразом их мог стать автокаталитический цикл, образованный теми самыми рибозимами, способными катализировать синтез своих собственных копий.
В.И. Вернадский о происхождении и сущности жизни и биосферы
В.И. Вернадский изложил свои взгляды о происхождении жизни в следующих тезисах:
- Начала жизни в том космосе, который мы наблюдаем, не было, поскольку не было начала этого космоса. Жизнь вечна, поскольку вечен космос, и всегда передавалась путем биогенеза.
- Жизнь, извечно присущая Вселенной, явилась новой на Земле, ее зародыши приносились извне постоянно, но укрепились на Земле лишь при благоприятных для этого возможностях.
- Жизнь на Земле была всегда. Время существования планеты — лишь время существования на ней жизни. Жизнь геологически (планетарно) вечна. Возраст планеты неопределим.
- Жизнь никогда не была чем-то случайным, ютящимся в каких-то отдельных оазисах. Она была распространена всюду, и всегда живое вещество существовало в образе биосферы.
- Древнейшие формы жизни — дробянки — способны выполнять все функции в биосфере. Значит, возможна биосфера, состоящая из одних прокариот. Вероятно, что такова она и была в прошлом.
- Живое вещество не могло произойти от косного. Между этими двумя состояниями вещества нет никаких промежуточных ступеней. Напротив, в результате воздействия жизни происходила эволюция земной коры.
Таким образом, необходимо признать, что к настоящему времени у современной науки нет однозначного ответа на вопрос о происхождении жизни.
Древнейший этап развития биосферы (после появления жизни)¶
После своего появления живые организмы благодаря присущей им биогеохимической функции начали преобразовывать, связанные в систему круговоротом воды, — литосферу, океан и атмосферу — в биосферу.
Продолжительность самого первого этапа развития биосферы оценивается в 2,5–2,9 млрд лет. Он занимал порядка 70–75 % всей истории биосферы и был в 2,5–4 раза продолжительнее добиосферного этапа развития нашей планеты.
Самым главным событием этого этапа было появление в океане в результате химической эволюции живого вещества, организованного в первые примитивные биоценозы. В их состав вошли цианобактерии и бактерии-органотрофы.
Геологическое время¶
Данный этап охватывает часть докембрийского эона, а именно архейский эон, следовавший за катархеем и часть протерозойского эона — палеопротерозойскую и мезопротерозойскую эры.
Временной промежуток с 4000 млн лет назад с границы между катархеем и археем до границы между мезопреторозойской и неопротерозойской эрами — 1000 млн лет назад (3000 млн лет).
Литосфера и тектоника¶
Временной период от 4100 до 3800 называют поздней тяжёлой бомбардировкой, во время которого на Луне, а также на Земле, Марсе, Венере, Меркурии образовались множество кратеров. Открытие поздней тяжёлой бомбардировки привело к пониманию того, какие процессы протекали ранее в катархейском эоне. До возникновения данной теории поверхность Земли в том периоде считалась покрытой лавой и огромными вулканами. Сейчас же поверхность Земли в тот период представляется твёрдой, покрытой кислой водой.
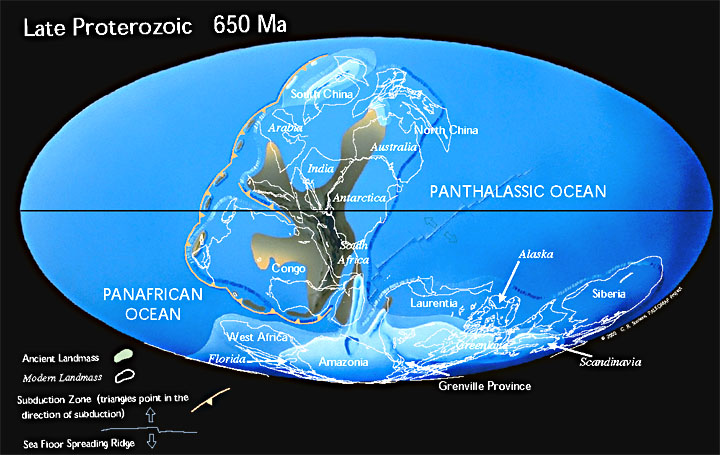
В мезоархейскую эру архейского эона около 3100 млн лет назад закончилось формирование первого гипотетического континента — Ваальбара, который существовал в период с 3600 до 2800 млн лет назад. В последствии он раскололся и сейчас части этой древней докембрийской платформы (кратона) (Kaapvaal и Pilbara) находятся в Южной Африке и в Западной Австралии.

В эту же эру формируется древнейший континент Ур, образовавшийся после Ваальбары примерно 3000 млн лет назад. В конце мезопротерозойской эры Ур объединился с континентами Нуна и Атлантика, сформировав суперконтинент Родиния. 300 млн лет назад он становится частью Пангеи, 208 млн лет назад Ур был разорван на части Лавразией и Гондваной. Участки земной коры, которые составляли Ур, теперь входят в состав Африки, Австралии и Индии.
2700 млн лет назад в неоархейской эре (конец архейского эона) формируется гипотетический суперконтинент Кенорленд (рис. ), является третьим по древности после Ваальбары и Ура.
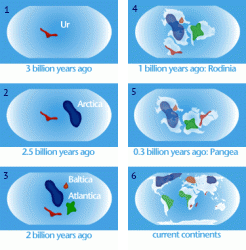
Начинается протерозойский эон и в палеопротерозойской эре формируется древний небольшой суперконтинент Нена, который состоял из кратонов Арктики, Балтики и Восточной Антарктиды. Континент был частью глобального суперконтинента Колумбия.
В период 1800–1500 млн лет назад формируется суперконтинент Колумбия (Нуна).

В конце мезопротерозойской эры формируется суперкинтинент Родиния, окруженный гигантским океаном Мировия. Возник около 1100 млн лет назад и распался около 750 млн лет назад. В то время Земля состояла из одной гигантской части суши и одного гигантского океана.
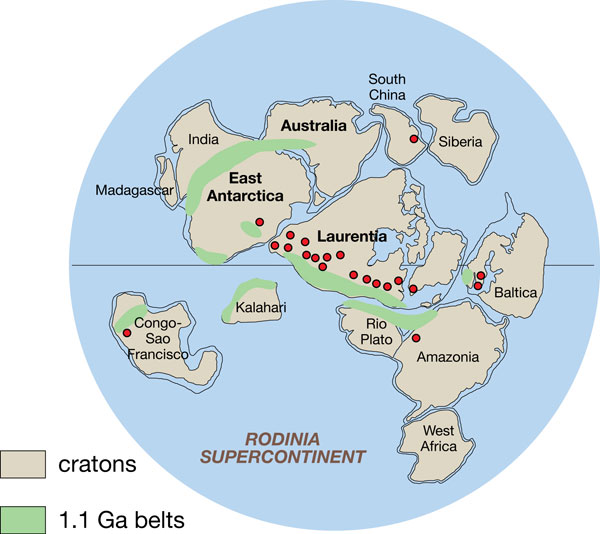
Геофизики предполагают, что до Родинии существовали и другие суперконтиненты: Кенорленд — максимальная сборка ≈ 2,75 млрд лет назад, Нуна (Колумбия) — максимальная сборка ≈1,8 млрд лет назад. После распада Родинии на Прото-Лавразию (северный континент) и Прото-Гондвану (южный континент) континенты вновь объединились в суперконтинент Паннотия ≈ 600 по 540 млн лет назад. После распада Паннотии континенты объединились в суперконтинент Пангея и снова распались в современное состояние.
Мировия — гипотетический всемирный океан, омывавший суперконтинент Родиния от 1100 до 800 миллионов лет назад, в эпоху неопротерозоя.
На данном этапе помимо базальтового слоя земной коры стал интенсивно формироваться и гранитный слой. Он сложил фундамент древних платформ, которые представляли собой ядра будущих континентов.
Также в этот период наблюдалось несколько эпох складчатости и магматической активизации (процесс повышения интенсивности тектонических движений и магматизма, обычно проявляющийся после периода относительного тектонического покоя).


Гидросфера¶
К концу древнейшего этапа океан занимал примерно 87 % поверхности Земли (против 98 % на предшествующем этапе). Зато был значительный (в 2,3 раза) прирост водной массы, которая достигла 80 % современного объёма океана. Почти в 3 раза увеличилась и средняя глубина океана (с 810 до 2510 м). Такое сокращение поверхности океана с одновременным увеличением его водной массы было следствием того, что рельеф земной поверхности стал более расчленённым, более контрастным, за счёт формирования древних платформ, которые по существу представляли собой ядра будущих континентов.
Появившиеся в океане древнейшие цианобактерии в водной среде океана образовывали на дне своеобразные маты. Их окаменевшие остатки, названные строматолитами, широко развиты во всех протерозойских породах. Именно строматолиты сформировали первые мощные карбонатные пласты на дне океана.
На протяжении этого этапа трижды наблюдался спад в развитии строматолитов, когда сокращались площади, занятые их постройками. Причину этих спадов исследователи видят в периодических изменениях климата, которые приводили к покровным оледенениям. Господствующие цианобактерии и бактерии органотрофы совместно освоили все биогеохимические функции живого вещества. Кислородную функцию исполняли цианобактерии, поскольку побочным продуктом их жизнедеятельности был кислород. Так в биосфере появился второй фотосинтетический источник свободного кислорода (наряду с эндогенным). Кислород из трофической системы стал поступать в океан, а затем и в атмосферу.
Усиление притока свободного кислорода за счёт фотосинтеза привело к существенным геохимическим последствиям. Так впервые под биогеохимическим воздействием живого вещества стал преобразовываться химизм естественных тел биосферы. Благодаря этому произошли кардинальные изменения в химизме океанических вод. До сих пор характер океанических вод был хлоридным с высоким содержанием восстановителей (сероводорода и закисного железа). Разросшаяся суша способствовала развитию на ней интенсивного химического выветривания. В результате с континентов в океан пошёл все возрастающий поток растворенных карбонатов натрия, калия, кальция, магния, железа и марганца. Это привело к тому, что хлоридный состав океанических вод преобразовывался и становился хлоридно-карбонатным.
Благодаря возросшему поступлению в океан кислорода за счёт фотосинтеза цианобактерий, постепенно окислялись такие восстановители, как сероводород и закисное железо. Они превращались в сульфаты и окислы железа. В результате океанические воды становились хлоридно-карбонатно-сульфатными.
После того, как были окислены все запасы сероводорода и восстановленного железа в водах первичного океана, кислород из океана стал поступать в атмосферу.
Масса воды в океане продолжала расти.
Атмосфера¶
Мы не знаем, какой была масса атмосферы на этом первом этапе развития биосферы, но, скорее всего, она была значительно меньше современной. Первоначальный состав атмосферы начал меняться за счёт появления кислорода.
- Кислород начал поступать в атмосферу сначала в результате фотохимических реакций с H2O и CO2. Но этот источник не мог дать более 0,1 % по сравнению с современной атмосферой.
- Вторым более мощным источником свободного кислорода был подводный базальтовый вулканизм, но первоначально поступивший из этого источника кислород шёл на окисление различных соединений, содержащихся в морской воде.
- И, наконец, третий источник (фотосинтез) — выделение свободного кислорода как побочного продукта жизнедеятельности цианобактерий.
Появление свободного O2 (сразу из трёх источников) привело к окислению CH4 → CO2 и, следовательно, к стабилизации карбонат-бикарбонатной системы в океанической воде. Углекислый газ (CO2) из атмосферы практически нацело был извлечён жизнью, и начали отлагаться биогенные карбонаты. Продукты серы — сероводород (H2S), оксид серы (SO2) и другие перешли в анион \(\ce{CO4^2–^}\), изменив состав океанической воды. Аммоний (катион \(\ce{NH4^1+^}\)), вероятно, был исчерпан организмами и окислен в элементарный азот (N). Так с помощью биоты создалась мощная современная азотно-кислородная атмосфера.
Как только содержание кислорода достигло примерно 1 % от современного, стала возможной жизнь аэробных организмов. Появление последних знаменует первую перестройку системы биосферы в истории её развития.
Климат¶
На этом этапе впервые фиксируются существенные изменения климата, которые выразились в трёх волнах резкого похолодания, которые сопровождались покровным оледенением.
В период палеопротерозойской эры около 2400 млн лет назад из-за большого количества поступающего кислорода метан, который ранее присутствовал в атмосфере в больших количествах и давал основной вклад в парниковый эффект, соединяется с кислородом и превращается в углекислый газ и воду, что приводит к значительному понижению общей температуры Земли. Изменения состава атмосферы, в свою очередь, привели к сокращению численности метаногенов, что вызвало дополнительное снижение уровня метана. Это событие названо кислородной катастрофой, которое привело к Гуронскому оледенению.

Гуронское оледенение является одним из древнейших и наиболее продолжительных оледенений на Земле. Началось и закончилось в палеопротерозое 2400–2100 млрд лет назад, продлившись около 300 млн лет. Геологические свидетельства оледенения лучше всего представлены в обнажениях горных пород к северу от озера Гурон в южной части Канады, в честь чего оледенение и получило своё название.

Биосфера¶
В период 4,1–3,8 млрд лет назад зарождается жизнь.
На этом древнейшем этапе развития биосферы главным событием было появление первых биоценозов живого вещества, образованных бактериями (цианобактериями и органотрофами). Именно благодаря их биогеохимической функции произошли серьёзные химические преобразования всех естественных тел биосферы. В целом в биосфере восстановительная среда постепенно сменялась окислительной.
Начало биологической эволюции (бактерии и сине-зелёные водоросли в океане). Начало фотосинтеза.
В эоархейскую эру архейского эона в период с 3900–3500 млн лет назад возникают клетки, похожие на прокариот. Первые организмы были хемотрофами и использовали углекислый газ как источник углерода, окисляли неорганические материалы, чтобы извлечь из них энергию. Затем они развивают гликолиз, набор химических реакций, высвобождающих энергию органических молекул, таких как глюкоза, и хранящих ее в химических связях АТФ. Гликолиз и сейчас используется почти всеми организмами.
В палеархейскую эру около 3500 млн лет назад происходит разделение протоорганизмов на бактерии и археи. Бактерии развивают примитивные формы фотосинтеза, которые по началу не производят кислород.
В ископаемых слоях отложений возраста 3400 млн лет появляются первые окаменелости микроорганизмов, метаболизм которых использовал серосодержащие соединения.
В конце архейского эона (3000–2700 млн лет назад) появляются фотосинтезирующие цианобактерии. Они используют воду как восстановитель, производя в результате кислород как отходы. В мезоархее уже встречаются строматолиты.
Концентрация кислорода в атмосфере существенно повышается, действуя как яд для многих видов бактерий. Луна всё ещё очень близко к Земле и вызывает приливы высотой до 300 метров, а поверхность постоянно терзается ураганными ветрами. Возможно, такие экстремальные условия смешивания значительно простимулировали эволюционные процессы.
В начале протерозойского эона (1850 млн лет назад) в отложениях появилась Grypania spiralis один из древнейших организмов. Возможно этот организм представлял собой гигантские бактерии или бактериальные колонии, однако огромные размеры (более 10 мм) и постоянство формы говорит о том, что окаменелости могли принадлежать эукариотическим водорослям, которые, таким образом, являются древнейшими известными эукариотическими организмами.

Наиболее древние останки этого вида относятся к породам возрастом 2,1 млрд лет железистой формации Негауни в штате Мичиган, где обнаружены сотни видимых невооружённым глазом спиралевидных углеродистых окаменелостей. Похожие окаменелости возрастом 1,1–1,4 млрд лет найдены в американском штате Монтана, Китае и Индии. Возможно, что Grypania является фотосинтезирующей водорослью.
Также около 1700 млн лет назад в палеопротерозойскую эру в палеонтологической летописи появляются клетки, содержащие ядро — эукариоты.
Эукариотическая клетка содержит выполняющие различные функции органеллы, которые окружены мембраной. По теории симбиогенеза некоторые органеллы, например митохондрии или хлоропласты (играющие роль «живых электростанций», производящих АТФ), произошли от прокариот путём симбиоза. Первоначально митохондрии были отдельными клеточными организмами, бактериями-друзьями, которые сосуществовали вместе с другими клетками и помогали осуществлять им некоторые функции. По прошествии некоторого времени они захвачены своими хозяевами, постепенно утратили способность к самостоятельному существованию и превратились в органоиды (органеллы). Переход клеток к производству энергии с использованием митохондрий стал эволюционной революцией, так как открыл путь к дальнейшему развитию ядерных клеток и усложнению их внутренней структуры.
В мезопротерозойскую эру около 1200 млн лет назад развиваются первые многоклеточные организмы, в основном состоящие из колоний клеток ограниченной сложности.
Появление в ископаемых слоях красных водорослей. У этих растений впервые возникает половое размножение, увеличив скорость эволюции. Появляются первые неморские эукариоты.
В конце мезопротерозойской эры 1060–760 млн лет назад появляются первые грибы.
Неопротерозойский этап развития биосферы¶
Неопротерозойский этап охватывает период 570–1000 млн лет назад. Этот этап длился до рубежа в 570 млн лет тому назад, когда живые организмы освоили кальциевую функцию, что привело к очередной перестройке биосферы. Таким образом, длительность рассматриваемого этапа примерно 430 млн лет или около 12 % продолжительности всей истории биосферы.
Геологическое время¶
Данный входит полностью в неопротерозойскую эру протерозойского эона и включает в себя три периода: Тонский, Криогенский, Эдиакарский в промежуток времени 1000–541 млн лет назад.
Литосфера и тектоника¶
Около 750 млн лет назад происходит раскол Родинии на Прото-Лавразию (северный континент), протоплатформу Конго и Прото-Гондвану (южный континент).
При смещении Прото-Лавразии к Южному полюсу, частичном повороте Прото-Гондваны и внедрении между ними Конголезской платформы приблизительно 600 миллионов лет тому назад образовалась Паннотия и просуществовала до 540 млн лет назад.
Продолжались эпохи складчатости и магматической активизации.
Продолжался рост гранитного слоя земной коры, который слагал древние платформы Гондванскую, Северо-Американскую, Восточно-Европейскую, Сибирскую и Китайскую. Этот континентальный блок уже занимал чуть меньше 25 % земной поверхности.
Гидросфера¶
Поверхность океана все еще продолжала сокращаться, при этом его водная масса увеличилась на 12 %. Она достигла 92 % от современного объёма. Увеличилась средняя глубина океана до 3200–3300 м, но среди морских бассейнов существовали как глубоководные, так и мелководные, расположенные на древних платформах.
Живое вещество по-прежнему было сосредоточено только в океане, в структуре которого произошли существенные изменения. Увеличение содержания кислорода в атмосфере до 1 % от современного привело к увеличению его концентрации и в морской воде, а это создало условия для жизни аэробных организмов.
Неопротерозойская революция происходила в океане, который по своему химическому составу начал приближаться к современному составу океанических вод.
В океане продолжалось хемогенное и биогенное образование больших толщ карбонатных пород — известняков и доломитов. Такое захоронение в осадках большого количества CO2 приводило к падению его содержания в атмосфере.
Атмосфера¶
Благодаря фотосинтезу зелёных водорослей атмосфера стала приобретать азотно-кислородный состав. Она освобождалась (благодаря интенсивному карбонатообразованию) от CO2 всё больше приближаясь к современной атмосфере, хотя концентрации отдельных газов в ней ещё отличались от современных. Содержание кислорода достигло 0,01 % от современного уровня.
Запасы атмосферного кислорода в период 580–540 млн лет позволили сформироваться озоновому слою, который блокировал ультрафиолетовое излучение, позволяя организмам выйти на сушу.
Климат¶
В неопротерозойской эре произошло несколько масштабных оледенений.
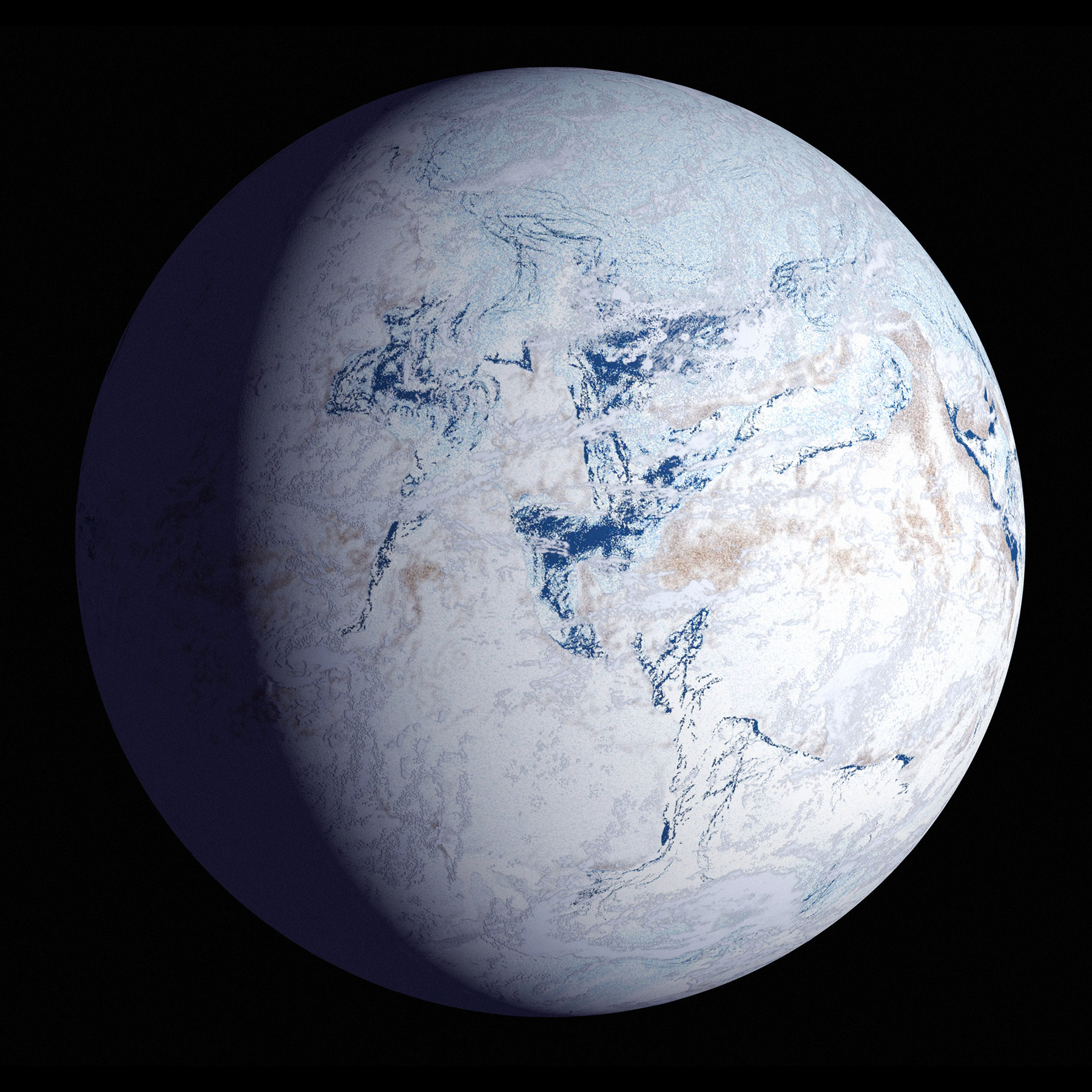
В период 850–630 млн лет (криогенский период неопротерозойской эры) произошло глобальное оледенение. Большая часть Родинии была расположена вокруг южного полюса, а окружавший её океан покрыт льдом толщиной в два километра. Лишь часть Родинии — будущая Гондвана — находилась вблизи экватора.
Существует гипотеза «Земля — снежный ком» (Snowball Earth), предполагающая, что Земля была полностью покрыта льдом в части криогенийского и эдиакарского периодов неопротерозойской эры, а также, возможно, в другие геологические эпохи.
Биосфера¶
Развитие многоклеточных организмов (многоклеточные водоросли, кораллы-археоцеаты). Первые организмы с нервной системой (черви).
Первая перестройка системы биосферы произошла около 1 млрд лет назад, в начале позднего протерозоя. Именно к этому рубежу приурочена неопротерозойская революция в развитии живого вещества, в результате которой широкое развитие получили эукариоты. Появилась та принципиально новая форма живого вещества, с жизнедеятельностью и биогеохимической функцией которого связано кардинальное преобразование всей биосферы. В первую очередь потоков энергии и круговоротов химических элементов.
На этом этапе продолжалось дальнейшее разрастание древних платформ, на которых формировались мелководные морские бассейны. В этих мелководных морях получили широкое распространение эукариоты, как одноклеточные, так и примитивные многоклеточные. Именно их широкое развитие стало главным событием этого этапа.
Появление фотосинтеза у эукариот резко усилило кислородную биогеохимическую функцию живого вещества и создало второй, фитосинтетический (в дополнение к эндогенному) этап поступления свободного кислорода. Это, в свою очередь, способствовало более быстрому геохимическому и биогеохимическому преобразованию водных и воздушных масс биосферы, которые по своему составу стали приближаться к современным.
Грандиозное оледенение должно было подавить растительную жизнь на Земле и, следовательно, привести к значительному снижению концентрации или даже полному исчезновению кислорода, что позволило образоваться неокисленным, богатым железом породам. Скептики утверждают, что такое оледенение должно было привести к полному исчезновению жизни, чего не произошло. Сторонники гипотезы отвечают им, что жизнь могла выжить следующими путями:
- Оазисы анаэробной и аноксифильной жизни, питаемые энергией глубоководных гидротерм, выживали в глубине океанов и коры — но фотосинтез был там невозможен.
- В открытом океане, вдалеке от суперконтинента Родиния или его осколков после его распада, могли остаться небольшие участки открытой воды, сохранившие жизнь с доступом света и углекислого газа для фотосинтетиков, дававших небольшие количества кислорода, достаточные для поддержания некоторых оксифильных организмов. Такой вариант возможен и в том случае, если океан замёрз полностью, но небольшие участки льда были достаточно тонки, чтобы пропускать свет.
- На нунатаках в тропиках, где днём тропическое солнце или вулканическое тепло разогревали скалы, защищённые от холодного ветра, и образовывали временные талые водоёмы, замерзавшие после заката.
- Споры и покоящиеся стадии, вмороженные в лёд, могли пережить наиболее тяжёлые фазы оледенения.
- Под слоем льда, в хемолитотрофных экосистемах, теоретически ожидаемых в ложах современных ледников, высокогорной и арктической вечной мерзлоте. Особенно вероятно это в зонах вулканизма или геотермальной активности.
- В бассейнах жидкой воды внутри и под слоем льда, подобно озеру Восток в Антарктиде. Согласно теории, эти экосистемы подобны микробным сообществам, живущим в постоянно замёрзших озёрах антарктических сухих долин.
Российский палеонтолог Михаил Федонкин, однако, указывая, что современные данные говорят о том, что большинство групп эукариотических организмов появилось ещё до неопротерозойского оледенения, считает это свидетельством против «крайних палеоклиматических моделей в виде гипотезы Земля-снежок, не отрицая роль похолодания в эукариотизации биосферы.
Многоклеточная жизнь, представленная в эдиакарском периоде 580–500 млн лет назад — причудливые, продолговатые, по большей части неподвижные, организмы, формой напоминающие лист. Ископаемые следы, оставленные по всему миру, раскрывают впервые появившуюся у многоклеточных организмов явную двустороннюю (билатеральную) симметрию.
Кроме симметрии, например, у сприггины (возможный предок членистоногих) **хорошо заметна наметившаяся «голова», образованная первыми двумя сегментами, и основное «тело», уменьшающееся к «хвосту». Появляется структура, которая будет повторяться у большинства сложных организмов.



Первое свидетельство о половом размножении у животных — фунизия, а также первые ископаемые свидетельства появления зубов, пищеварительного тракта и ануса у маркуэлиа.



В период с 580 по 540 млн лет назад появляются гребневики — самые большие среди передвигающихся при помощи ресничек животных.

Первые ископаемые свидетельства морских губок и коралловых полипов (кораллы и актинии).





Раннепалеозойский этап развития биосферы¶
Геологическое время¶
Закончился докембрийский эон и начался фанерозойский эон. Данный этап охватывает часть часть палеозойской эры фанерозойского эона — кембрийский, ордовикский и силурийский периоды примерно с 541 по 419 млн лет назад. Продлился он около 120–170 млн лет.
По своей продолжительности он был сравнительно коротким (охватил всего лишь около 4–4,8 % всей длительности истории биосферы), но значимость его была колоссальна, так как в течение его создались условия, которые позволили живому веществу освоить новое для них пространство — сушу возникших континентов.
Литосфера и тектоника¶
В кембрии, всего через 60 млн лет после своего образования, Паннотия распалась на континент Гондвана и мини-континенты Балтики, Сибири и Лаврентии. Материки были затоплены мелководными морями. Суперконтинент Гондвана только что сформировавшийся находился недалеко от Южного полюса.
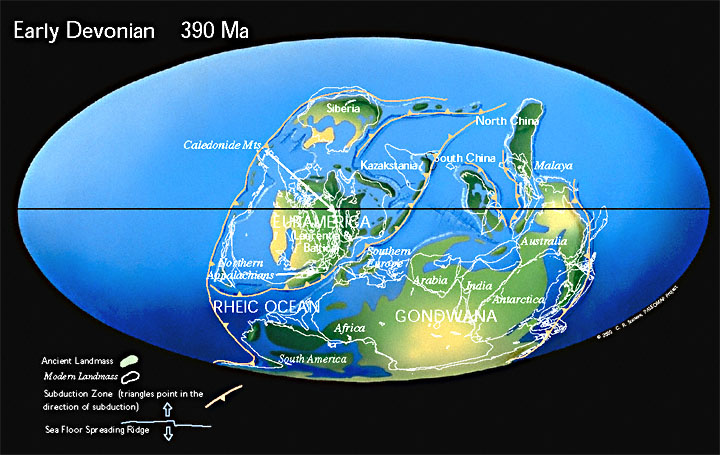
Период Каледонской складчатости (каледонский орогенез), наиболее ранние фазы которой относятся к середине — концу кембрия, основные фазы захватывают конец ордовика — начало силура.
В ордовикском периоде древние океаны Панталасса, Япетус и Палеотетис разделяют континенты Лаврентию, Балтику, Сибирь и Гондвану. Конец ордовика был одним из самых холодных времен в истории Земли. Лед покрыл большую часть южной части Гондваны.
В силурийском периоде континенты начинают сталкиваться с закрытием палеозойских океанов. Лаврентия сталкивается с Балтикой, закрывая северную ветвь океана Япетус и образуя континент «Древний красный песчаник». Коралловые рифы начинаю занимать всё большие площади и наземные растения начинают колонизировать бесплодные континенты.
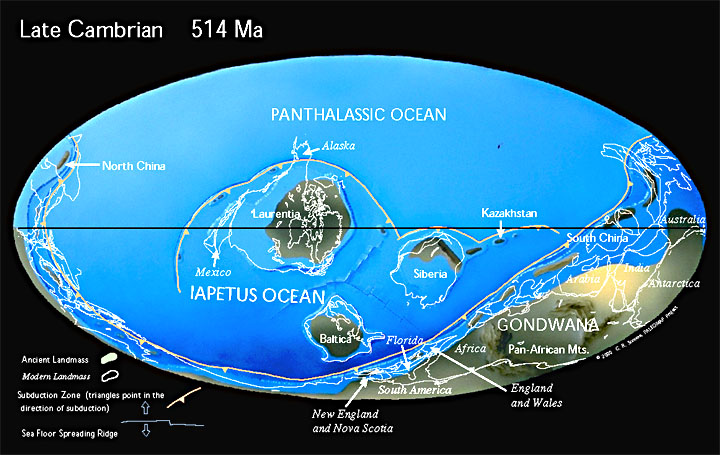
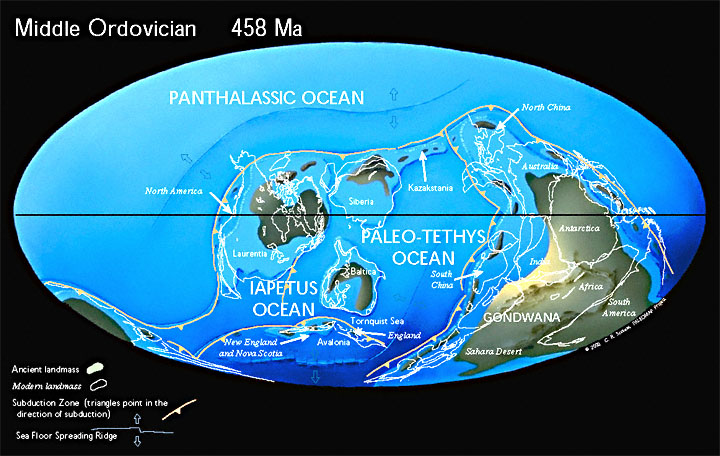
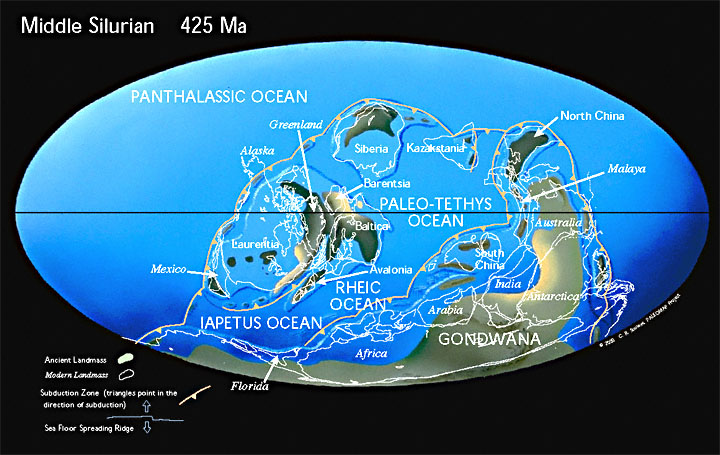
Разросшиеся в предшествующее время блоки гранитного слоя земной коры образовали древние платформы со сравнительно ровным рельефом. Платформы, ядра будущих континентов, были существенно приподняты над океаническим дном, расположенным на базальтовом слое земной коры. Этот континентальный блок занимал порядка 23 % земной поверхности.
Гидросфера¶
Океан на этом этапе занимал около 77 % поверхности земли. Но характер морских бассейнов претерпел существенные изменения. Среди них выделялись:
- мелководные эпиконтинентальные моря (в пределах шельфа материка),
- геосинклинальные бассейны с более расчленённым рельефом,
- и глубоководные океанические бассейны.
Временами морские воды заливали древние платформы, полностью или частично. Такие наступления моря называются трансгрессиями. В результате образовывались мелководные обширные эпиконтинентальные моря, где кипела жизнь. Временами воды уходили с древних платформ, которые вновь становились сушей. Такой отход моря называют регрессиями морского бассейна. Для рассматриваемого этапа было характерно три волны трансгрессий и регрессий. По окраинам древних платформ и между ними развивались более глубоководные геосинклинальные бассейны с резко расчленённым рельефом дна и развитым вулканизмом.
Главным сосредоточением морских организмов были мелководные платформенные моря и геосинклинальные бассейны.
Интенсивное извлечение карбоната кальция из морской воды привело к тому, что морские воды из хлоридно-карбонатно-сульфатных становились хлоридно-сульфатными, приближаясь по своему химическому составу к водам современного океана. Однако их солёность не оставалась постоянной. Она то увеличивалась, то уменьшалась. Разросшиеся континентальные массивы суши поставляли все больше и больше растворимых солей в океан, что не могло не вызвать увеличение его солёности.
Атмосфера¶
Как уже говорилось, благодаря фотосинтетической деятельности водорослей, которые получили широкое распространение в мелководных бассейнах, шло постепенное накопление кислорода в атмосфере. Одновременно сокращалось в ней и содержание CO2. Он использовался живыми организмами для построения внутреннего или наружного карбонатного скелета.
К концу данного этапа содержание кислорода в атмосфере приблизилось к современному.
Климат¶
В ордовикском и силурийском периоде около от 450 млн до 420 млн млн лет назад произошло небольшое Андо-Сахарское оледенение, продлившееся около 30 миллионов лет.
Биосфера¶
Органический мир кембрия, ордовика и силура за последние 200 лет достаточно хорошо изучен палеонтологами. Долгое время исследователей поражала внезапность появления с самого раннего кембрия всех основных типов животного мира и низших растений. Но потом было выяснено, что эта внезапность только кажущаяся. Она объясняется тем, что живые организмы с начала кембрия освоили кальциевую функцию и стали строить свои раковины и скелеты из карбоната кальция. Это способствовало лучшему сохранению их остатков в горных породах. Поэтому, хотя все основные типы животных и примитивных низших растений появились ещё во время неопротерозойской революции, на предыдущем этапе, но они смогли продемонстрировать все своё разнообразие только тогда, когда у них появились твёрдые скелеты и раковины.
Среди животных преобладали беспозвоночные, хотя в ордовике уже появились первые бесчелюстные позвоночные. Все животные и низшие растения обитали преимущественно в мелководных морях, приуроченных к древним платформам. К концу этого этапа биоразнообразие выросло примерно в 10 раз.
Возможно, что на этом этапе бактериальные сообщества начали осваивать подземную биосферу.
В связи с тем, что среди эукариотов (растений и животных) появились новые формы, которые освоили построение минерального скелета из карбоната кальция стал меняться газовый состав атмосферы: в ней уменьшилось содержание углекислого газа и увеличилось содержание кислорода за счёт фотосинтеза водорослей, которые получили массовое развитие в мелководных морях.
Кембрийский взрыв видообразования
В период 540–500 млн лет назад происходит кембрийский взрыв — относительно быстрое (всего за несколько миллионов лет) появление в палеонтологической летописи большей части современных биологических типов, сопровождаемое сильным увеличением видового разнообразия у других, включая животных, фитопланктон и кальцимикробов.
Кембрийским взрывом называют явление, результатом которого стало резкое увеличение количества ископаемых остатков живых существ в отложениях, датируемых началом кембрийского периода, то есть около 540 млн лет назад. Именно в этот период появляются в палеонтологической летописи останки хордовых, членистоногих, моллюсков и иглокожих.
Фактором кембрийского взрыва была скелетная революция — появление у животных экзоскелета. В кембрии принципы взаимоотношений хищник-жертва распространились повсеместно, что и стало причиной возникновения минерализованных наружных скелетов. Начался новый виток эволюции, связанный с выработкой приспособлений к хищничеству и защите от хищников, в частности значительному развитию подверглись органы чувств (появляются совершенные глаза и мозг).
Проблематика «кембрийского взрыва» состоит в попытках объяснить внезапное появление окаменелостей на нижней границе кембрия и их отсутствие в более древних отложениях — как свидетельствах существования или несуществования жизни в предшествующий период истории Земли. В кембрийском периоде основная жизнь была сосредоточена в морях. Каковы причины кембрийского взрыва?
Первая группа возможных причин: - изменение окружающей среды: - рост концентрации кислорода. Самая ранняя атмосфера Земли вообще не содержала свободного кислорода. Тот кислород, которым дышат современные животные — как содержащийся в воздухе, так и растворенный в воде — является продуктом миллиардов лет фотосинтеза, главным образом — микроорганизмов. Примерно 2,5 миллиарда лет назад концентрация кислорода в атмосфере резко возросла. До этого времени весь вырабатываемый микроорганизмами кислород полностью тратился на окисление элементов с высоким сродством к кислороду, таких как железо. Пока не произошло их полное связывание на суше и в верхних слоях океана, в атмосфере существовали лишь локальные «кислородные оазисы». Нехватка кислорода могла длительное время препятствовать развитию крупных сложных организмов. Проблема состоит в том, что количество кислорода, которое животное может абсорбировать из окружающей среды, ограничено площадью поверхности (лёгких и жабр у наиболее сложных животных; кожи — у более простых). Количество же кислорода, требуемое для жизнедеятельности, определяется массой и объёмом организма, которые по мере увеличения размеров растут быстрее, чем площадь. Рост концентрации кислорода в воздухе и в воде мог ослаблять или вовсе устранять это ограничение. - земля-снежок. Существуют многочисленные доказательства того, что в позднем неопротерозое Земля подвергалась глобальному оледенению, в ходе которого большая часть её была покрыта льдом, а температура поверхности была близка к точке замерзания даже на экваторе. Некоторые исследователи указывают, что это обстоятельство может быть тесно связано с кембрийским взрывом, поскольку самые ранние из известных ископаемых относятся к периоду вскоре после конца последнего полного оледенения.
Вторая группа возможных причин: - анатомические объяснения: - развитие полового размножения. Возможное появление полового размножения или его существенное развитие в период кембрийского взрыва для очень примитивных и похожих существ может означать то, что существовала возможность их межвидового и более далёкого скрещивания. Это резко увеличивало изменчивость. Лишь с развитием генома появляются по-настоящему изолированные виды, не скрещивающиеся с другими.
Третья группа возможных причин: - экологические объяснения. Такие объяснения сосредоточены на взаимодействии между различными видами организмов: - «гонка вооружений» между хищниками и жертвами. Хищничество по определению предполагает гибель жертвы, в силу чего оно становится сильнейшим фактором и ускорителем естественного отбора. Давление на жертв в направлении лучшей адаптации должно быть более сильным, чем на хищников — поскольку, в отличие от жертвы, хищник имеет шанс сделать новую попытку (эта асимметрия известна как принцип «жизнь против обеда» — хищник рискует потерей только обеда, в то время как жертва рискует жизнью). - множество пустых ниш. Резкие изменения в строении тела являются «затруднительными»; изменения имеют гораздо больше шансов на существование, если они встречают слабую (или вовсе не встречают) конкуренцию за ту экологическую нишу, на которую они нацелены. Последнее необходимо, чтобы новый тип организмов имел достаточно времени для адаптации к своей новой роли. Это обстоятельство должно приводить к тому, что реализация основных эволюционных изменений гораздо более вероятна на начальных стадиях формирования экосистемы — из-за того, что последующая диверсификация заполняет почти все экологические ниши. В дальнейшем, несмотря на то, что новые типы организмов продолжают возникать, нехватка пустых ниш препятствует их распространению в экосистеме. Эта модель хорошо объясняет факт уникальности кембрийского взрыва — почему он случился только один раз и почему его длительность была ограничена.
Происходит сильная диверсификация живых существ в океанах: хордовые, членистоногие (например трилобиты и ракообразные), иглокожие, моллюски, плеченогие, фораминиферы, радиолярии и другие.
Потребовалось 3 млрд лет для появления многоклеточных организмов, но всего 70–80 млн лет для того, чтобы скорость эволюции возросла на порядок и породила основную часть сегодняшнего видового разнообразия.
В начале кембрийского периода (примерно 540 млн лет назад) у некоторых групп животных появляется сложно устроенный глаз.
Также в кембрии около 530 млн лет назад появляются первые ископаемые отпечатки следов на земле, которые указывают на то, что ранние животные исследовали сушу ещё до того, как на ней появились растения.
510 млн лет назад появляются первые головоногие (наутилоидеи) и панцирные моллюски.
485 млн лет назад появляются первые позвоночные с настоящими костями (бесчелюстные). В морях позднего кембрийского периода основными группами животных были членистоногие, иглокожие и моллюски. Но самым важным обитателем морей того времени было бесчелюстное существо хайкоуихтис — у него кроме глаз развилась хорда.
450 млн лет назад на суше появляются двупарноногие (класс многоножек), а в море — конодонты и морские ежи.
Ордовикско-силурийское вымирание
443,7 млн лет назад. Вымерло более 60 % морских беспозвоночных, включая две трети семейств брахиопод и мшанок. Причинами катастрофы могли быть вулканизм и эрозия или вспышка гамма-излучения от сверхновой звезды.
Массовое вымирание на границе ордовикского и силурийского периода, около 450–440 миллионов лет назад. Третье по процентной части вымерших родов из пяти сильнейших вымираний в истории Земли и второе — по потерям в количестве живых организмов.
В период между 450 и 440 млн лет назад, с промежутком в 1 млн лет, произошли два всплеска вымирания. Для морских организмов это вымирание второе по величине, оно уступает только пермскому вымиранию. В то время вся известная жизнь обитала в морях и океанах. Погибло более 60 % морских беспозвоночных, включая две трети всех семейств брахиопод и мшанок. Особенно пострадали брахиоподы, двустворчатые моллюски, иглокожие, мшанки и кораллы.
Непосредственной причиной вымирания, как представляется, явилось движение Гондваны к области южного полюса. Это привело к глобальному похолоданию, оледенению и последовавшему за ним падению уровня мирового океана. Отступление границы океана разрушило или повредило места обитания вдоль континентального побережья. Данные об оледенении были найдены в отложениях в пустыне Сахара. Комбинация низкого уровня мирового океана, похолодания и образования ледников, вероятно, и послужила причиной ордовикского вымирания.
440 млн лет назад появляются первые представители групп бесчелюстных — гетеростраки и галеаспиды.
434 млн лет назад первые примитивные растения «выходят» на сушу, развившись из зелёных водорослей. Растения сопровождали грибы, которые могли помогать им завоёвывать сушу с помощью симбиоза. Рстение, к примеру, куксонии (Coocsonia), которые достигали в высоту не более 10 см, и некоторые виды лишайников.
428 млн лет назад — первое ископаемое свидетельство сухопутного членистоногого. У некоторых членистоногих развились примитивные лёгкие, позволявшие им дышать атмосферным воздухом, например, скорпион бронтоскорпио мог находиться на суше в течение четырёх часов.
420 млн лет назад появляются ранние лучепёрые рыбы, панцирные пауки и сухопутные скорпионы. Первые гигантские грибы прототакситы, достигавшие 8,8 метра в высоту.
В силурийских морях появляются окончательно сформировавшиеся позвоночные животные. В отличие от членистоногих, у позвоночных имелся костный хребет, позволявший им лучше маневрировать под водой. У позвоночного цефаласписа, например, также развились сенсорные органы, которые генерировали особое магнитное поле, позволявшее ему ощущать окружающую среду. Также у цефаласписа развился примитивный мозг, позволявший животному запоминать некоторые события.



Позднепалеозойский этап развития биосферы¶
Геологическое время¶
Данный этап охватывает конец палеозойской эры — девонский, каменноугольный и пермский периоды начало и мезозойской эры — триасовый период.
Длился данный промежуток времени с 419 по 201 млн лет назад.
Очередная перестройка биосферы произошла около 410–400 млн лет назад, в конце силура, когда образовался наземный растительный покров, изменивший все биогеохимические и геофизические процессы. Именно тогда произошло формирование четвёртого естественного тела биосферы глобальной размерности — ландшафтной оболочки, появившейся на континентах и островах.
Именно это привело к окончательному формированию той принципиальной схемы потоков энергии в биосфере, которая существует до настоящего времени. Все естественные тела глобальной размерности стали приобретать современный облик, в том числе и круговороты химических элементов. В наземных условиях открылись новые возможности для эволюционных преобразований живого вещества. И они были реализованы появлением в мире растений — голосемянных, а в животном царстве — среди беспозвоночных — насекомых, а среди позвоночных — их наземных форм.
Литосфера и тектоника¶
В девоне ранние палеозойские океаны закрылись, образуя «допангею». Пресноводные рыбы смогли мигрировать с континентов южного полушария в Северную Америку и Европу. Впервые леса выросли в экваториальных районах арктической Канады.
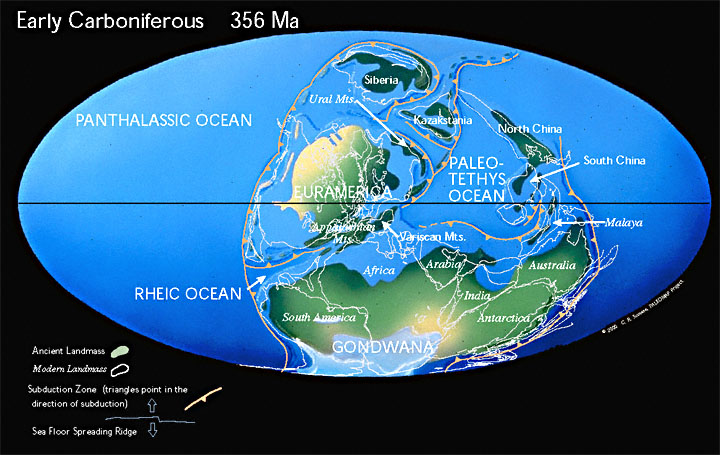
В раннем карбоне начинает формироваться Пангея. В раннем карбоне палеозойские океаны между Еврамерикой (Лавруссия) и Гондваной начали смыкаться, образуя Аппалачи и горы Варискан. На Южном полюсе росла ледяная шапка.
Пангея — сверхконтинент, существовавший в конце палеозоя и начале мезозоя и объединявший практически всю сушу Земли 335–175 млн лет назад — Гондвану, Лаврентию, Балтию и Сибирь.
В процессе формирования Пангеи из более древних континентов на местах их столкновения возникли горные системы. Некоторые из них просуществовали и до нашего времени, к примеру Урал или Аппалачи. Эти горы гораздо древнее таких сравнительно молодых горных систем, как Альпы в Европе, Кордильеры в Северной Америке, Анды в Южной Америке или Гималаи в Азии. Из-за длящейся много миллионов лет эрозии Урал и Аппалачи сильно разрушены и невысоки.
Гигантский океан, омывавший Пангею, носит название Панталасса. Панталасса — гипотетический океан, окружавший, начиная с силурийского периода (443 млн лет назад) палеозоя и до среднего мезозоя (200 млн лет назад) включительно, суперконтинент Пангею и покрывавший около половины земного шара. В настоящее время литосферные плиты Панталассы замещены литосферными плитами под Тихим океаном.
Огромный залив, называемый морем Тетис, вдавался в суперконтинент между будущими Евразией и Австралией.
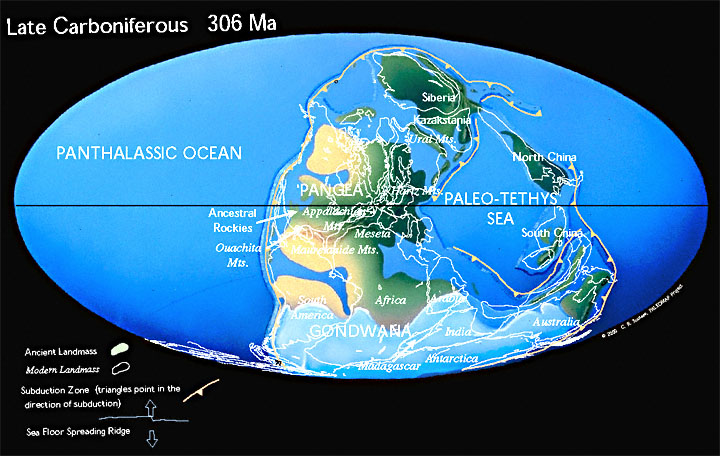
В позднем карбоне континенты, которые составляют современную Северную Америку и Европу, столкнулись с южными континентами Гондваны, образуя западную половину Пангеи. Лед покрыл большую часть южного полушария, а вдоль экватора образовались обширные угольные болота.
В конце пермского периода было величайшее вымирание за все время. Обширные пустыни покрывали западную Пангею во время пермского периода, когда рептилии распространились по поверхности суперконтинента. 99 % всего живого погибло во время вымирания, которое ознаменовало конец палеозойской эры.
В конце триаса Пангея начала раскалываться (раскололась в Юрском периоде). Суперконтинент Пангея, в основном собранный в триасовый период, позволил наземным животным мигрировать с Южного полюса на Северный полюс. После великого пермско-триасового вымирания и потепления Тетису биоразнообразие начало снова восстанавливаться.
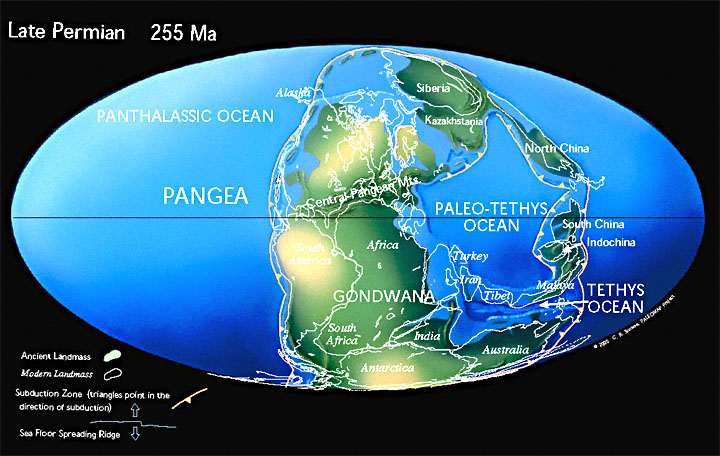
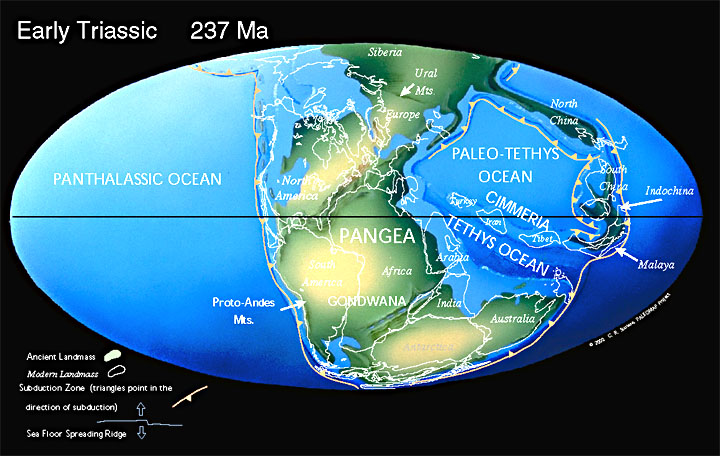
Земная кора континентального типа продолжала наращиваться, как вокруг древних платформ, так и путём образования молодых платформ (Западно-Европейской, Марокканской, Скифско-Туркменской).
Периодически (через 40–60 млн лет) возникали орогенные эпохи, когда создавались горы. Это сопровождалось резким усилением наземного вулканизма. Во время вулканических извержений увеличивалось поступление на дневную поверхность радиоактивных элементов, что в свою очередь вело к повышению естественного радиоактивного фона как наземной среды, так и морской воды.
Гидросфера¶
Океан на этом этапе больше приобретал свои современные черты, хотя соотношение суши и моря, а также объем океанических вод не оставались постоянные. На протяжении этапа наблюдалось четыре морских трансгрессии, во время которых морские воды проникали далеко в глубь материков, образуя обширные мелководные эпиконтинентальные моря. В них, равно как и в геосинклинальных морях и открытом океане, обитали самые разнообразные морские животные и растения. Состав их видов, родов и семейств не оставался постоянным, он постоянно обновлялся. Наиболее крупные изменения произошли на рубеже палеозоя и мезозоя.
Усложнение дна океана. Заложение котловины Тихого океана. Объем воды в океане приблизился к современному.
Ландшафтная сфера¶
Начало формирования самого молодого, по меркам геологического времени, естественного тела биосферы глобальной размерности — ландшафтной сферы — было положено выходом растений и животных на сушу в конце силура–начале девона.
Такое расширение поля жизни стало возможным после того, как возник последний защитный уровень биосферы — озоновый экран. После его возникновения первыми сушу заселили микроорганизмы: силикатные бактерии и лишайники. Они приняли самое активное участие в процессах выветривания горных пород. И с этого момента выветривание превратилось из физико-химического процесса в биогеохимический. Заселение новой сухопутной ниши микроорганизмами привело также к уменьшению содержания CO2 в атмосфере и гидросфере.
В ландшафтной оболочке с самого начала возникает ряд весьма существенных новообразований, которые сыграли значительную роль в развитии всей биосферы.
Первый такой феномен — почвы, в которых содержание живого вещества достигает нескольких десятков процентов массы. Первым шагом к образованию почв было превращение микроорганизмами процесса выветривания из физикохимического в биогеохимический. Вторым шагом было развитие в карбоне сплошного покрова наземной растительности, которая своими корнями скрепила образовавшийся на материнских породах верхний рыхлый слой.
Второй феномен — болота и связанное с ними угленакопление. Одновременно с образованием наземного растительного покрова появились и болота, в которых началось угленакопление, достигшее своего максимума в карбоне и начале перми. Потом эпохи угленакопления регулярно повторялись при периодически наступавших увлажнениях климата. Обилие влага способствовало пышному развитию наземных растений.
Преобразования, происшедшие среди наземных живых организмов, были значительны. Ландшафтная оболочка — совершенно другая среда обитания. Только на суше живые организмы в полной мере испытали на себе силу земного тяготения. В водной среде они развивались по сути в невесомости. На суше и совсем другая симметрия среды, нежели в водной толще океана. Поэтому в ландшафтной сфере у живых организмов появилось много новых возможностей для развития в самых разных направлениях.
Атмосфера¶
Атмосфера постепенно становится все более азотно-кислородной.
В девоне содержание кислорода приблизилось к современному. Концентрация углекислого газа изменилась от 0,1 до 0,4 %. Тропический влажный, умеренно влажный и сухой климат. К концу периода — гумидизация климата.
В карбоне преобладает гумидный климат. Средняя температура 26,6 °С в начале и 21 °С в конце. В конце периода — оледенение на всех континентах Южного полушария.
В перми химический состав, строение и циркуляция близки к современным. Континентальный засушливый климат. Планетарное похолодание (понижение температуры на 3–4 градуса). Оледенение в Гондване и локальное оледенение в Северном полушарии.
В триасе установился современный тип циркуляции атмосферы. Разноообразие и зональность (от тропического до холодного) климатов. Преобладание аридности.
Климат¶
Оледенение Карру
360 млн лет назад в девоне начинается оледенение Карру, продлившееся примерно 100 до 260 млн лет назад.
Биосфера¶
Надо отметить, что в самом начале этапа, в девонском периоде, в морях были широко представлены самые разнообразные рыбы — панцирные, кистеперые, двоякодышащие и так далее. Недаром девонский период называют «веком рыб». От некоторых из них произошли первые наземные позвоночные.
410 млн лет назад появляются первые признаки образования зубов у рыб. Самые ранние плауновидные.
395 млн лет назад появляются первые лишайники и харовые водоросли (ближайшие родственники наземных растений). Ранние сенокосцы, клещи, ногохвостки и аммониты.
В отложениях возраста 375 млн лет обнаружена тиктаалик — лопастепёрая рыба, живущая в мелких реках, болотах или озёрах. Стала переходным звеном между рыбами и земноводными, обладая рёбрами, схожими с теми, что есть у четвероногих; подвижным шейным отделом и примитивными лёгкими, которые позволяли ей недолго находиться на суше.


Пышно разросшиеся листопадные растения сбрасывают свою листву в тёплые и бедные кислородом водоёмы, привлекая тем самым мелкую добычу и затрудняя обитание там больших хищных рыб. Исследователи полагают, что тиктаалик, скорее всего, развил свои прото-конечности, передвигаясь по дну и иногда выползая на берег на короткое время.
Время жизни древнейшей из известных живородящих организмов — панцирной рыбы Materpiscis. Она вынашивает потомство в своём организме. Эта адаптация позволяет защитить плод от агрессивной среды в критический период развития нового организма и снабдить его питательными веществами через пуповину.

II. Девонское вымирание
374 млн лет назад. Уничтожает около 19 % семейств и 50 % родов. Это вымирание было одним из крупнейших в истории земной флоры и фауны. Исчезают почти все бесчелюстные. Представляло собой серию эпизодов сокращения численности таксонов на протяжении практически всего среднего, верхнего и даже частично нижнего девона. Даже фоновый уровень вымирания девонского периода составлял 18,8 % родов.
Массовое вымирание видов в конце девона, одно из крупнейших в истории Земли вымираний флоры и фауны. Первый (и самый сильный) пик вымирания произошёл около 374 млн лет назад, когда неожиданно исчезли почти все бесчелюстные. Второй импульс завершил девонский период (около 359 млн лет назад). Всего вымерло 19 % семейств и 50 % родов.
К позднему девону суша была полностью освоена и заселена растениями, насекомыми и земноводными, а моря и океаны были полны рыбы. Кроме того, в этот период уже существовали гигантские рифы, сформированные кораллами и строматопоратами. Евро-американский континент и Гондвана только начали движение друг к другу, чтобы в будущем сформировать суперконтинент Пангею. Вероятно, вымирание в основном затронуло морскую жизнь.
Основная теория предполагает, что главной причиной вымирания в океанах послужили изменения в уровне океана и обеднение океанических вод кислородом. Возможно, что активатором этих событий послужило глобальное похолодание или обширный океанический вулканизм, хотя падение внеземного тела, такого как комета, тоже вполне возможно.
Так как «вымирание» происходило на протяжении длительного периода, то очень трудно выделить единственную причину, повлёкшую вымирание и даже отделить причину от следствия.
363 млн лет назад к началу каменноугольного периода Земля начинает походить на современную. Насекомые уже ползают по суше и скоро они покорят небо; в океанах плавают акулы, а растения, распространяющие семена, уже покрыли поверхность суши и уже скоро вырастут и разрастутся первые леса.
Четвероногие (тетраподы) приспосабливаются к изменившемуся миру и заселяют сушу. Они постепенно утрачивают признаки, свойственные своим предкам — кистепёрым рыбам, такие как жабры и чешуя и, приспосабливаясь к жизни на суше, начинают дышать только лёгкими. Их голова становится ещё подвижнее, чем у тиктаалика из-за более развитого шейного отдела, а конечности набирают силу и подвижность. Эти существа потом разделятся на 4 класса: земноводные, пресмыкающиеся, птицы и млекопитающие.
360 млн лет назад — первые крабы и папоротниковые. На земле доминируют семенные папоротники.
350 млн лет назад — первые крупные акулы, химеровые и миксиновые.
330 млн лет назад — первые позвоночные-амниоты (имеющие зародышевые оболочки), например, палеотирис (небольшая рептилия).
312 млн лет назад датируется древнейший известный отпечаток тела насекомого, предка подёнки.
305 млн лет назад — самые ранние рептилии-диапсиды (например, петролакозавры).

300 млн лет назад — количество кислорода в атмосфере достигает 30–35 % (сейчас 20 %), это позволяет некоторым насекомым, таким как меганевра, достигать поистине гигантских размеров. Размах её крыла достигал 75 см. Это самое большое известное науке летающее насекомое, вместе с пермской Meganeuropsis permiana.
280 млн лет назад — появляются первые жуки. Растёт разнообразие семенных и хвойных деревьев, в то время как лепидодендралии и сфенопсиды постепенно вымирают. Увеличивается видовое разнообразие земноводных (темноспондильные) и пеликозавров. В океанах появляются первые геликоприоны.
III. Пермское вымирание
Датируется 252,2 млн лет назад.
Является самым массовым вымиранием из всех, приведшее к исчезновению 57 % всех семейств и 83 % всех родов, более 90 % (по некоторым оценкам, даже 95 %) морских видов (наибольшие потери зафиксированы для фузулинид, ругоз, ракушковых и различных классов плеченогих; также пострадали мшанки, гастроподы, губки и сидячие двустворчатые) и более 70 % видов наземных позвоночных.
При этом поражающие механизмы действовали избирательно: среди сидячих организмов с массивным карбонатным скелетом вымерло до 90 % родов, для обладателей менее массивных скелетов, в зависимости от возможности метаболически контролировать диффузионный газообмен, этот показатель составлял от 28 % до 78 %, а для таксонов с лёгким или вообще отсутствующим карбонатным скелетом — всего 10 %; также показано, что более сильному вымиранию подверглись таксоны, характеризовавшиеся бо́льшим (на начало кризиса) разнообразием и более высоким уровнем фонового вымирания. Примечательно, что пермская катастрофа произошла за очень короткий по геологическим меркам промежуток времени — менее 200 тыс. лет.
На восстановление биоразнообразия наземных организмов понадобилось 50 млн лет, а морских — до 100 млн лет. Все экологические связи были разрушены и впоследствии выстраивались заново.
По нему проведена граница между пермским и триасовым геологическими периодами (палеозойской и мезозойской эр).
В настоящее время у специалистов отсутствует общепринятое мнение о причинах вымирания. Рассматривается ряд возможных причин: - катастрофические события: усиление вулканической деятельности в Сибири; падение одного или многих метеоритов, либо столкновение Земли с астероидом диаметром в несколько десятков километров (одним из доказательств этой гипотезы служит возможное наличие 500-километрового кратера в районе Земли Уилкса — часть территории Восточной Антарктиды); внезапный выброс метана со дна моря; приобретение археями способности перерабатывать органику с выделением больших объёмов метана. - постепенные изменения окружающей среды: изменения химического состава морской воды и атмосферы, в частности, дефицит кислорода; повышение сухости климата; изменение океанических течений и/или уровня моря под влиянием изменений климата. Наиболее распространена гипотеза, согласно которой причиной катастрофы явилось излияние траппов (особый тип континентального магматизма, для которого характерен огромный объём излияния базальта за геологически короткое время на больших территориях), которое могло повлечь за собой вулканическую зиму, парниковый эффект из-за выброса вулканических газов и другие климатические изменения, повлиявшие на биосферу. Считается, что на восстановление биосферы после массового вымирания ушло около 30 млн лет, однако некоторые учёные делают выводы, что оно могло произойти и за более короткий промежуток времени, около 5–10 млн лет. Вымирание старых форм открыло дорогу многим животным, долгое время остававшимся в тени: начало и середина следующего за пермью триасового периода ознаменовалось становлением архозавров, от которых произошли динозавры и крокодилы, а впоследствии птицы. Кроме того, именно в триасе появляются первые млекопитающие.
225 млн лет назад появляются самые ранние динозавры (прозавроподы). Питаются растениями и становятся первыми крупными динозаврами, появившимися на Земле. Первые сердцевидки, видообразование у саговниковидных, беннеттитовых и хвойных. Первые костистые рыбы.

220 млн лет назад леса голосеменных доминируют на суше; травоядные достигают гигантских размеров. Большой размер даёт им лучшую защиту от хищников и позволяет иметь длинный кишечник, необходимый для лучшего переваривания растений, бедных питательными веществами. Первые двукрылые и черепахи (одонтохелис). Первые динозавры-целофизоиды.
215 млн лет назад появляются первые млекопитающие (например эозостродон). Небольшое количество видов позвоночных вымирает.
IV. Триасовое вымирание
208–200 млн лет назад — триасовое вымирание, — в результате которого вымерло 23 % всех семейств и 48 % всех родов, по меньшей мере, половина известных сейчас видов, живших на Земле в то время, в частности, 20 % морских видов (в наибольшей степени — двустворчатые, конодонты, известковые губки, каменистые кораллы, подклассы головоногих наутилоиды и аммоноиды), некоторые растения, множество пресмыкающихся, а также последние крупные земноводные, что, освободило экологическую нишу для динозавров.
Были предложены несколько объяснений этого события: - Постепенная перемена климата или флуктуации уровня океана в течение позднего триасового периода. Однако, это не объясняет внезапность вымирания существ в океане. Возможно, перемена климата связана с изменением площади мирового океана и его глубины, вызванной движением земной коры. Отражение солнечного света водной поверхностью и увеличение влажности климата могло спровоцировать появление полярных и высокогорных ледяных шапок, что привело в свою очередь к ледниковому периоду и значительному промерзанию акватории океана. - Падение астероида. Однако ударный кратер не обнаружен, время образования которого совпадало бы с триасово-юрской границей. - Массовые извержения вулканов, особенно излияние базальтовых лав в Центрально-атлантической магматической области, высвободило бы в атмосферу углекислый газ или диоксид серы, которые, в свою очередь, явились бы причиной сильного глобального потепления (от первого газа) или похолодания (от второго газа). - Гипотеза о метангидратном ружье. Потепление из-за вулканизма и накопления углекислого газа в атмосфере, могло привести к высвобождению метана из донных отложений. Выделение метана, даже более сильного парникового газа чем CO2, ускорило бы потепление ещё сильнее, что, в свою очередь, привело бы к ещё большему высвобождению метана со дна океанов. Этот процесс мог привести к быстрому изменению глобальной температуры.
Исчезает по меньшей мере половина известных на сегодняшний день видов, живших на Земле в то время. Это событие освобождает экологические ниши и позволяет динозаврам начать доминировать на суше. Триасовое вымирание прошло менее чем за 10000 лет, непосредственно перед началом распада Пангеи.






Современный этап развития биосферы¶
Геологическое время¶
Современный этап развития биосферы начался с той перестройки, которую она испытала около 150 млн лет назад, когда, установились те константы геохимических равновесий, что сохранились до настоящего времени. Этот последний этап по своей продолжительности самый короткий: он охватывает всего около 4 % времени существования биосферы с 201 млн лет назад по настоящее время.
Он включает в себя два периода мезозойской эры — юрский и меловой и полностью кайнозойскую эру с тремя периодами — палеогеновый, неогеновый и четвертичный.
Литосфера и тектоника¶
Юра
В начале Юрского периода около 200 млн лет назад Пангея начинает распадаться на Лавразию и Гондвану, разделённые океаном Тетис. Оба суперконтинента далее будут распадаться на более мелкие части и их столкновения приведут к активному горообразованию. Результатом давления Африки (отколющейся от Гондваны) на Европу (часть Лавразии) станут Альпы, а столкновение Индии (Гондвана) и Азии (Лавразия) создаст Гималаи.
Лавразия (Лаврентия и Евразия) — древний континент, северный из двух континентов (южный — Гондвана), на которые распался протоконтинент Пангея в мезозое. Позже, от 135 до 200 млн лет назад, Лавразия распалась на Евразию и Северную Америку.
Лавразия включала большую часть территорий, которые ныне составляют континенты Северного Полушария. В палеозое эти территории входили в состав таких континентов, как Лавруссия (образовавшаяся из Лаврентии и Балтики), Сибирия, Казахстания, Северокитайский и Южнокитайский континентальные щиты.
Гондвана в палеогеографии — древний суперконтинент в южном полушарии, включавший Африку, Южную Америку, Антарктиду, Австралию, Новую Зеландию, а также Аравию, Мадагаскар и Индостан. Образовалась в конце докембрия (750–530 млн лет назад) в результате раскола суперконтинента Родиния.
Тетис — древний океан, существовавший в эпоху мезозоя между древними континентами Гондвана и Лавразия. Реликтами этого океана являются современные Средиземное, Чёрное и Каспийское моря.
Пангея начинает раскол. Суперконтинент Пангея начал распадаться в средней юре. В поздней юре Центральная часть Атлантического океана была узким океаном, отделявшим Африку от востока Северной Америки. Восточная Гондвана начала отделяться от Западной Гондваны.
150 млн лет назад в поздней Юре Гондвана распадается на две части, одна из которых включала Африку и Южную Америку, другая — Австралию, Антарктиду и полуостров Индостан.
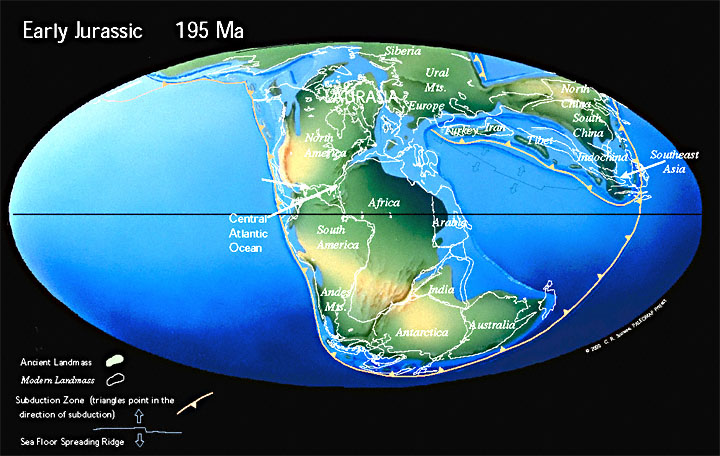
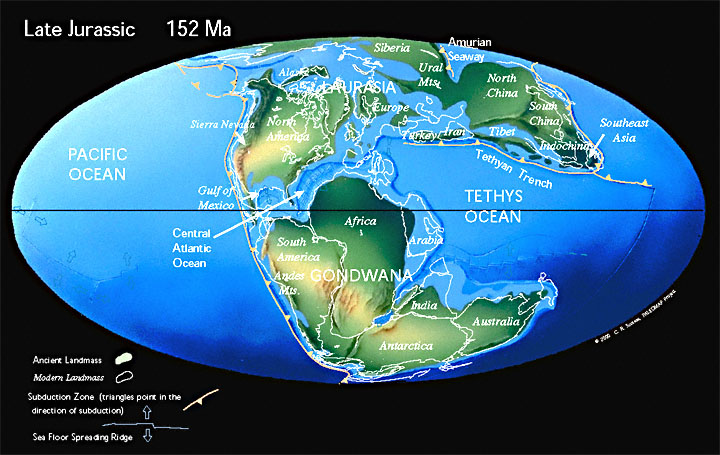
Мел
В меловой период открылся Южный Атлантический океан. Индия отделилась от Мадагаскара и устремилась на север, столкнувшись с Евразией. При этом Северная Америка была связана с Европой, а Австралия все еще была присоединена к Антарктиде.
55 млн лет назад Лавразия окончательно распадается на Лаврентию (сейчас Северная Америка) и Евразию (включая Индию).
Лаврентия — континент, существовавший в палеозойскую эру в восточной и центральной Канаде. Начал формирование 540 млн лет назад при распаде Паннотии и 335 млн лет назад объединился с другими континентами в Пангею.
Евразия — крупнейший из шести материков на Земле.
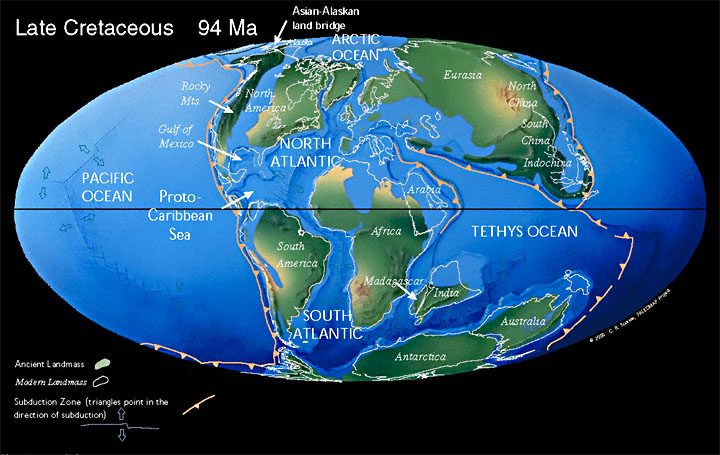
Мел-палеогеновая граница. Вымирают динозавры. Столкновение кометы шириной 10 км вызвало глобальные изменения климата, в результате которых погибли динозавры и многие другие формы жизни. К концу мелового периода океаны расширились, и Индия приблизилась к южной окраине Азии.
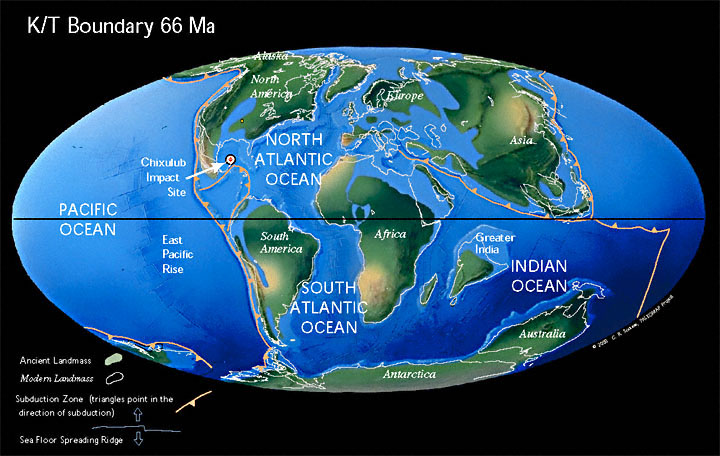
Эоцен
В раннем кайнозое Индия начала сталкиваться с Азией. 50–55 млн лет назад Индия начала сталкиваться с Азией, образуя Тибетское плато и Гималаи. Присоединенная к Антарктиде Австралия начала стремительно продвигаться на север.
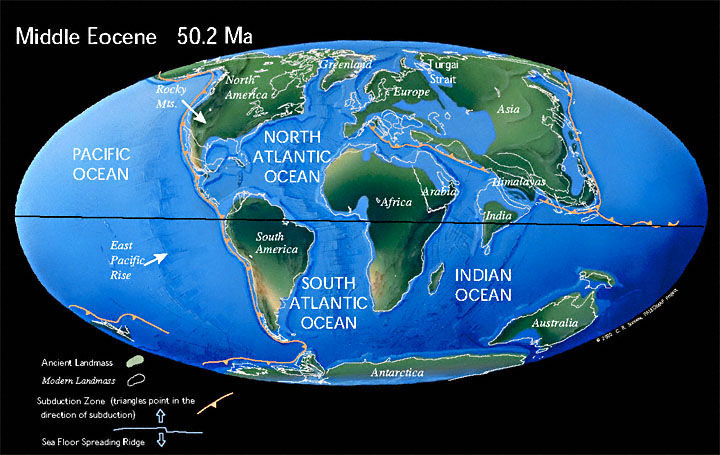
Миоцен
Мир принимает современную конфигурацию. 20 млн лет назад Антарктида была покрыта льдом, и северные континенты быстро охлаждались. Мир приобрел «современный» вид, но обратите внимание, что Флорида и некоторые части Азии были затоплены морем.
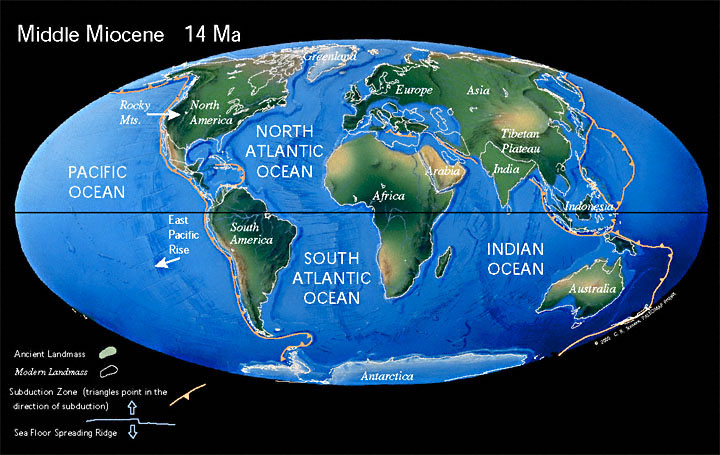
Максимум последнего оледенения — заключительный этап последней ледниковой эпохи. Земля находилась в климате ледяного дома последние 30 млн лет. Когда Земля находится в климатическом режиме «Ледяной дом», на полюсах есть лед. Полярный ледяной покров расширяется и сжимается из-за изменений орбиты Земли (циклы Миланковича). Последнее расширение полярных ледяных щитов произошло около 18000 лет назад.
Современный мир имеет четко определенные климатические зоны. Мы вступаем в новую фазу столкновения континентов, которая в конечном итоге приведет к образованию нового суперконтинента Пангея в будущем. Глобальный климат нагревается, потому что мы выходим из ледникового периода и потому, что мы добавляем в атмосферу парниковые газы.
Гидросфера¶
Охлаждение поверхностных и глубинных вод. Понижение уровня Мирового океана в конце периода.
Атмосфера¶
В Юре произошло выравнивание климатических условий, усилился тепло- и влагообмен. Теплый относительно влажный климат, средняя температура воздуха 24,5 °С.
В мелу климат оставался влажным, средняя температура 25,2 °С. Похолодание в середине и конце периода.
В палеогене теплый и умеренно влажный климат. Средняя температура 22–23 °С. Уменьшение контрастности температур между тропическими и полярными областями. Похолодание в конце периода.
В неогене похолодание и аридизация климата. Оледенение в Антарктиде и Северном полушарии (в конце периода).
В антропогене оледенение в Северном полушарии.
Биосфера¶
Главная особенность этапа состояла в том, что к его началу внутренняя структура биосферы окончательно сложилась: все естественные тела глобальной размерности уже сформировались и приобрели современные очертания, сложилась и система потоков энергии в биосфере, а также круговоротов химических элементов.
В начале этапа под влиянием внешних причин скачкообразно изменились основные геохимические константы. Это, вероятно, вывело систему биосферы из сложившегося ранее неустойчивого равновесия, чем создало условия для возникновения новообразований среди живых организмов. Поэтому для этого этапа основные события связаны с крупными эволюционными преобразованиями живого вещества, приуроченного к ландшафтной сфере. Строение и состав атмосферы, океана и земной коры литосферы быт весьма близок к современным параметрам.
В Юрском периоде около 200 млн лет назад появляются вирусы. В это же время произошли крупные вымирания среди наземных позвоночных, в частности, крупных земноводных. Появляются самые ранние виды анкилозавров.
195 млн лет назад появляются первые птерозавры-доригнатусы и динозавры-зауроподы. Увеличение видового разнообразия маленьких птицетазовых динозавров: гетеродонтозаврид, фаброзаврид и сцелидозаврид.



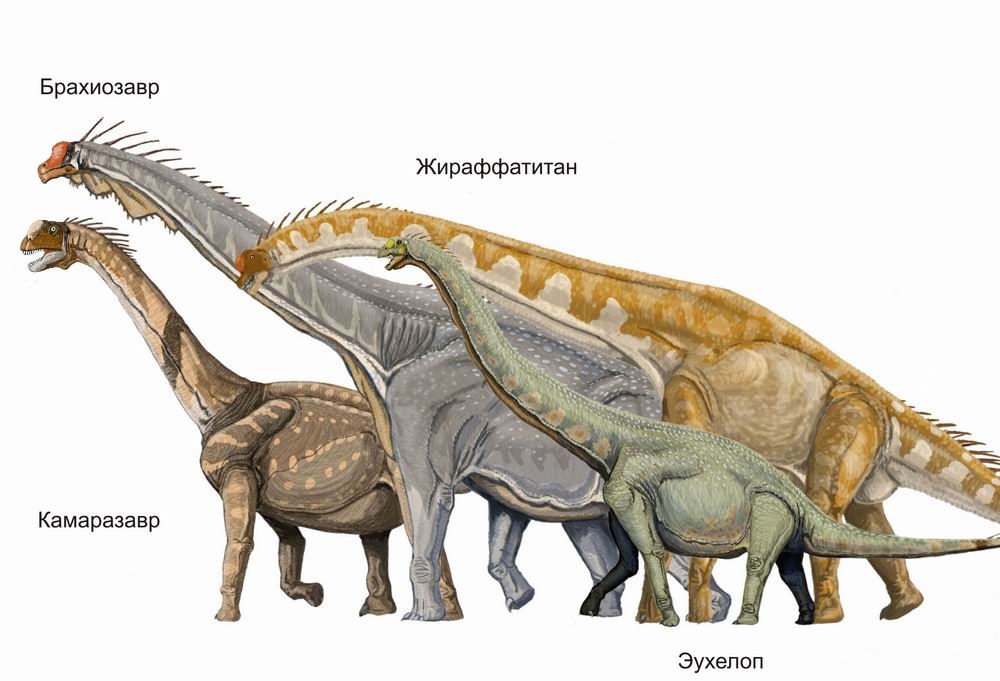
В окаменелостях, датируемыми 190 млн лет, появляются плиозавры. Первые бабочки (археолепис), раки-отшельники, современные морские звёзды, неправильные морские ежи, двустворчатые корбулиды и мшанки (tubulipore bryozoans). Обширное образование губковых рифов.


176 млн лет назад — первые стегозавры.


170 млн лет назад — первые саламандры, тритоны, криптоклидиды и эласмозавриды (плезиозавры), и млекопитающие кладотерии. Цинодонты вымерли, в то время как видов зауропод стало больше.
165 млн лет назад — первые скаты и двустворчатые глицимеридиды.
161 млн лет назад — в палеонтологической летописи появляются травоядные цератопсы.


160 млн лет назад — первое плацентарное млекопитающие Juramaia sinensis, предок всех высших зверей и человека, живёт на территории будущей провинции Ляонин.


155 млн лет назад — первые кровососущие насекомые (мокрецы), рудистовые двустворчатые и мшанки. Археоптерикс, одна из первых птиц, появляется в окаменелостях, вместе с млекопитающими триконодонтидами и симметродонтами. Увеличивается разнообразие у стегозавриев.



Птицы среди сухопутных позвоночных животных появились около 150 млн лет назад. Это событие было весьма значимым для эволюции биосферы. Птицы, наряду с насекомыми, освоили для своего передвижения воздушную среду и стали играть важную роль в миграции химических элементов в биосфере.


В меловом периоде 140 млн лет назад — появляются первые покрытосеменные.
130 млн лет назад — рост разнообразия покрытосеменных (цветковых) растений: они развивают специальные структуры, привлекающие насекомых и других животных, чтобы с их помощью обеспечивать опыление. Такая инновация вызвала бурное эволюционное развитие через коэволюцию.
К тому моменту, когда в меловое время появились цветковые растения, насекомые уже прочно освоились в биосфере, распространившись по всему пространству ландшафтной оболочки. Именно эта их «всюдность» и помогла им сыграть решающую роль в возникновении и дальнейшем прогрессе доминирующих в настоящее время цветковых растений. Как считал академик А.Л. Тахтаджан, травянистые формы покрытосеменных произошли от их древесных форм. Современная цветковая растительность, насчитывающая 390 семейств, около 13000 родов и не менее 240000 видов, составляет основную массу растительного вещества биосферы. А без насекомых она существовать не может. Иными словами, насекомые являются самыми непосредственные участниками производства основной первичной продукции биосферы. Хотя опыление цветковых растений и стало важнейшей функцией насекомых, этим далеко не исчерпывается их биосферная роль. Не менее важно, что они ещё и регуляторы численности беспозвоночных, а также важнейший источник белковой пищи для самых разных животных. Ныне классу насекомых принадлежит более 70 % видов животного царства. Насекомые активно включились в трансформацию органического вещества в биосфере.
Первые пресноводные пеломедузовые черепахи.
115 млн лет назад — первые однопроходные млекопитающие.
110 млн лет назад — первые гесперорнисообразные и зубастые ныряющие птицы. Самые ранние двустворки из семейств лимопсиды, вертикордииды и тиазириды.
106 млн лет назад — появление спинозавра, самого крупного тероподного динозавра.
100 млн лет назад — первые пчёлы. Ископаемый род мелитосфекс считается «вымершей ветвью собирателей пыльцы из надсемейства Apoidea, дочерней к современным пчёлам», и датируется нижним мелом.
90 млн лет назад — вымирание ихтиозавров. Самые ранние змеи и двустворчатые нукуланиды. Сильная диверсификация у покрытосеменных: магнолид, розид, гамамелисовых, однодольных и имбиря. Первые известные клещи.
80 млн лет назад — первые муравьи и термиты.
70 млн лет назад — увеличения видового разнообразия у многобугорчатых млекопитающих. Первые двустворки-йолдииды.
68 млн лет назад — тираннозавр, самый крупный наземный хищник Северной Америки, появляется в ископаемых слоях. Первые виды трицератопсов.


66 млн лет назад — около полуострова Юкатан падает 10-километровый астероид. Удар с энергией 100 тератонн в тротиловом эквиваленте создаёт 180-километровый кратер Чикшулуб и вызывает цунами высотой 50–100 метров. Помимо очевидных катастрофических последствий в виде ударной волны и цунами, это столкновение выбросило в атмосферу на значительную высоту много пыли и серы. Эти частички могли оседать около года, что уменьшило в этот период количество солнечной энергии, достигающей земной поверхности, на 10–20 %. Есть предположения, что удар пришёлся в крупный резервуар с нефтью, из-за чего она, попав в воздух, взорвалась, что объясняет наличие крошечных углеродных сфер диаметром порядка 50 мкм в породах этого периода.
Существуют гипотезы, что данное падение было лишь одним из нескольких, на что указывает наличие кратера Шива и Болтышского кратера на территории Украины. Падение крупного тела рядом с Индией могло вызвать вулканические извержения расположенных поблизости траппов Декана. Примерно в ту же эпоху и возникает мощный вулканизм в Индии, что сильно и очень быстро изменяет климат Земли и ставит динозавров на грань гибели.
V. Мел-палеогеновое вымирание
65,5 млн лет назад. Уничтожает около половины всех видов животных, включая мозазавров, птерозавров, плезиозавров, аммонитов, белемнитов, рудистовых и иноцерамидовых двустворчатых, большую часть планктонных фораминифер и всех динозавров, кроме их потомков — птиц.
Произошло на границе мелового и палеогенового периода (граница мезозойской и кайнозойской эр).
Вместе с динозаврами вымерли морские рептилии, в том числе мозазавры и плезиозавры, летающие ящеры (птерозавры), многие моллюски, в том числе аммониты и белемниты, и множество мелких водорослей. Всего погибло 16 % семейств морских животных (47 % родов морских животных) и 18 % семейств сухопутных позвоночных, включая практически всех крупных и средних по размеру. Все существовавшие в мезозое экосистемы были полностью разрушены, что впоследствии резко подстегнуло эволюцию таких групп животных, как птицы и млекопитающие, давших в начале палеогена огромное многообразие форм благодаря освобождению большинства экологических ниш.
Причины вымирания: - Гипотеза импакта. Падение астероида — одна из самых распространённых версий. Она основана главным образом на приблизительном соответствии времени образования кратера Чикшулуб на полуострове Юкатан в Мексике и временем вымирания большинства из исчезнувших видов динозавров. - Версия «многократного падения». Она привлекается, в частности, для объяснения того, что вымирание произошло не одномоментно. - Усиление вулканической активности, с которой связывают ряд эффектов, которые могли бы повлиять на биосферу: изменение газового состава атмосферы; парниковый эффект, вызванный выбросом углекислого газа при извержениях; изменение освещённости Земли из-за выбросов вулканического пепла (вулканическая зима). В пользу этой гипотезы говорят геологические свидетельства о гигантском излиянии магмы в промежутке между 68 и 60 млн лет назад на территории Индостана, в результате которого образовались деканские траппы. - Резкое понижение уровня моря, произошедшее в последней фазе мелового периода. - Резкий скачок магнитного поля Земли. - Переизбыток кислорода в атмосфере Земли. - Резкое охлаждение океана. - Эпизоотия — массовая эпидемия у животных. - Динозавров истребили первые хищные млекопитающие, уничтожая кладки яиц и детёнышей. - Постепенное изменение климата, вызванное дрейфом материков.
Ни одна из перечисленных гипотез не может в полной мере объяснить весь комплекс явлений, связанных с вымиранием динозавров и других видов в конце мелового периода.

В Палеогеновый период 65 млн лет назад начинается быстрое распространение хвойных и гинкговых в высоких широтах, вместе с млекопитающими, становящимися доминантным классом. Первые псаммобииды. Быстрое увеличение количества видов муравьёв.
Пургаториус, маленький предок плезиодапиморфов, успешно переживает глобальную катастрофу и становится первым прото-приматом — наиболее вероятным предшественником всех приматов. Наш наиболее вероятный предок был всего 10 сантиметров в длину и массов 20 г, жил на земле, активно передвигался и вероятнее всего рыл норы.
На Земле появились предки многих современных групп зверей — копытных, слонов, грызунов, приматов, рукокрылых (например, летучих мышей), китов, сирен. Мало-помалу млекопитающие начинают покорять земной шар.
63 млн лет назад — эволюционирование креодонтов, важной группы плотоядных млекопитающих.
60 млн лет назад 3 диверсификация больших нелетающих птиц. Появляются первые настоящие приматы, вместе с первыми двустворками-семелидами, неполнозубыми, хищными и насекомоядными млекопитающими и совами. Предки плотоядных млекопитающих (миацид) становятся многочисленными.
55 млн лет назад — повышается разнообразие групп современных птиц (первые певчие птицы, попугаи, гагары, стрижи, дятлы), первый кит (гималайацетус), самые ранние грызуны, зайцы, броненосцы, появление сирен, хоботных, непарнокопытных и парнокопытных млекопитающих в ископаемых останках. Увеличивается разнообразие цветковых растений. В водных просторах плавает один из ранних представителей сельдевых акул, древняя акула-мако.
52 млн лет назад — появляются первые летучие мыши (онихониктерис).
50 млн лет назад — вершина разнообразия динофлагеллят и микроокаменелостей, рост разнообразия у фоладомиид и двустворчатых гетерокон. В слоях окаменелостей появляются бронтотериевые, тапиры, носороговые и верблюды. Увеличение разнообразия приматов.
40 млн лет назад — возникают современные формы бабочек и молей. Вымирание гасторнисов. Базилозавр, один из первых гигантских китов, появляется в окаменелостях.
35 млн лет назад — злаки. Лёгкое увеличение в разнообразии у хладостойких ракушковых и фораминифер, вместе с обширными вымираниями брюхоногих (улиток), пресмыкающихся и земноводных. Начинают возникать многие группы современных млекопитающих: первые глиптодонты, гигантские ленивцы, собаки, пекариевые, а также первые орлы и соколы. Разнообразие у зубатых и усатых китов.
33,9 млн лет назад — начинается малое Эоценово-олигоценовое вымирание, которое уничтожает около 3,2 % морских животных.
30 млн лет назад — первые усоногие и эвкалипты, вымирание эмбритоподовых и бронтотериевых млекопитающих, самые ранние кабаны и кошки.
28 млн лет назад — в отсутствие динозавров, как подавляющего фактора, млекопитающие быстро увеличивают свой размер — за первые 35 миллионов лет от мел-палеогенового вымирания размер видов увеличивался экспоненциально.
Появляется индрикотерий, самое большое сухопутное млекопитающее, когда-либо жившее на земле. Самые крупные особи достигали 8 м в высоту, а самые тяжёлые весили 20 тонн.
25 млн лет назад — первые олени.

В Неогеновый период 20 млн лет назад — появляются первые жирафы и гигантские муравьеды, увеличение разнообразия у птиц.
15 млн лет назад — в палеонтологической летописи появляются мастодонты, полорогие и кенгуру, увеличение разнообразия австралийской мегафауны.
10 млн лет назад — луга и саванны прочно заняли своё место на земле. Увеличение разнообразия насекомых, в особенности муравьёв и термитов. У лошадей увеличиваются размеры тела и развиваются передние верхние зубы. Сильное увеличение разнообразия у луговых млекопитающих и змей.
6 млн лет назад — диверсификация у австралопитековых.
5 млн лет назад — первые древесные ленивцы и бегемоты, разнообразие у луговых травоядных, больших плотоядных млекопитающих, норных грызунов, кенгуру, птиц и малых плотоядных. Стервятники набирают в размерах, уменьшение количества непарнокопытных млекопитающих. Вымирание плотоядных нимравид.
4,8 млн лет назад — мамонты появляются в ископаемых слоях.
4 млн лет назад — эволюция австралопитеков. Появляется ступендемис, становясь самой большой пресноводной черепахой.
3 млн лет назад — великий межамериканский обмен, когда различные наземные и пресноводные фауны мигрируют между Северной и Южной Америкой. Броненосцы, опоссумы, колибри и вампировые летучие мыши заселяют Северную Америку, в то время как тапиры, саблезубые кошки и олени мигрируют в Южную Америку. Появляются первые короткомордые медведи (арктодусы).
2,8 млн лет назад — появляется Человек умелый (Homo habilis). Происходит диверсификация хвойных в высоких широтах. В Индии появляется вероятный предок крупного рогатого скота — тур.

В Четвертичный период 1,7 млн лет назад — вымирают австралопитековые.
1,6 млн лет назад — дипротодон, крупнейшее известное сумчатое, когда-либо обитавшее на земле, появляется в ископаемых слоях. Этот представитель австралийской мегафауны просуществовал примерно полтора миллиона лет и вымер около 40 000 до н. э.
1,2 млн лет назад — эволюция Homo antecessor. Вымирают последние популяции парантропов.
600 тыс. лет назад — эволюция Homo heidelbergensis.
350 тыс. лет назад — эволюция неандертальцев.
300 тыс. лет назад — в Азии вымирают гигантопитеки, гигантские родственники орангутанов.
200 тыс. лет назад — в Африке появляется анатомически современный человек. Около 50000 лет назад он начал колонизацию других континентов, замещая неандертальцев в Европе и других гоминин в Азии.
73,5 тыс. лет назад — население земли снижается примерно до 10000 (или даже до 1000) пар, что создаёт в эволюции человека эффект бутылочного горлышка, при этом сокращение генетического разнообразия приводит к изменению относительных и абсолютных частот аллелей генов, поэтому данный эффект рассматривается в числе факторов эволюции.
41 тыс. лет назад — денисовский человек живёт в большой пещере на территории, населённой также неандертальцами и современными людьми.
40 тыс. лет назад — вымирают последние известные гигантские вараны (мегалании).
30 тыс. лет назад — вымирание неандертальца.
20 тыс. лет назад — объём мозга у людей достигает максимума — 1500 см3 (сейчас 1350).
15 тыс. лет назад — последний из шерстистых носорогов умирает.
11 тыс. лет назад — наступает эпоха голоцена, сразу за последним ледовым максимумом. Гигантские короткомордые медведи (арктодусы) исчезают из Северной Америки вместе с последними гигантскими ленивцами. В Северной Америке вымирают все лошадиные.
10 тыс. лет назад — последние материковые популяции шерстистого мамонта вымирают, как и последние смилодоны.
6 тыс. лет назад — маленькие популяции американских мастодонтов вымирают в областях Юты и Мичигана.
4,5 тыс. лет назад — последние особи карликового подвида шерстистого мамонта исчезают с острова Врангеля.
393 года назад вымирают последние туры.
84 года назад последний сумчатый волк умирает в Тасманийском зоопарке 7 сентября 1936 года.


Организмы в биосфере¶
Видовое разнообразие организмов разного размера¶
Традиционно считалось, что чем мельче организмы какой-то группы, тем большим числом видов эта группа представлена. Однако оставалось неясным, как оценивать видовое разнообразие бактерий. Строгие правила научной номенклатуры предписывают, что бинарное латинское название (род, вид) может присваиваться только тем микроорганизмам, которые могут быть выделены в лаборатории в чистую культуру. Но таких видов сравнительно немного.
Чистая культура — совокупность микроорганизмов одного вида, имеющих одинаковые морфологические и биохимические свойства и одинаковые свойства их культур.
Относительно генетического разнообразия в пределах даже строго описанных видов бактерий в научном сообществе идут споры, считать ли это разнообразие внутривидовым или уже межвидовым. Поскольку среди ученых нет согласия относительно критериев вида у прокариот, приходится ограничиваться рассмотрением эукариот, тем более что они демонстрируют огромный диапазон размеров: если судить по длине тела, то от 10 мкм (некоторые инфузории) до десятков метров (китообразные), то есть в миллион раз.
Анализ связи между размерами тела и числом видов, проведенный для наземных организмов, показал, что с уменьшением размера тела число видов увеличивается, достигая максимума при размере тела около 1 см, а затем снижается. В тропических лесах большинство насекомых как раз имеют размер около 1 см.
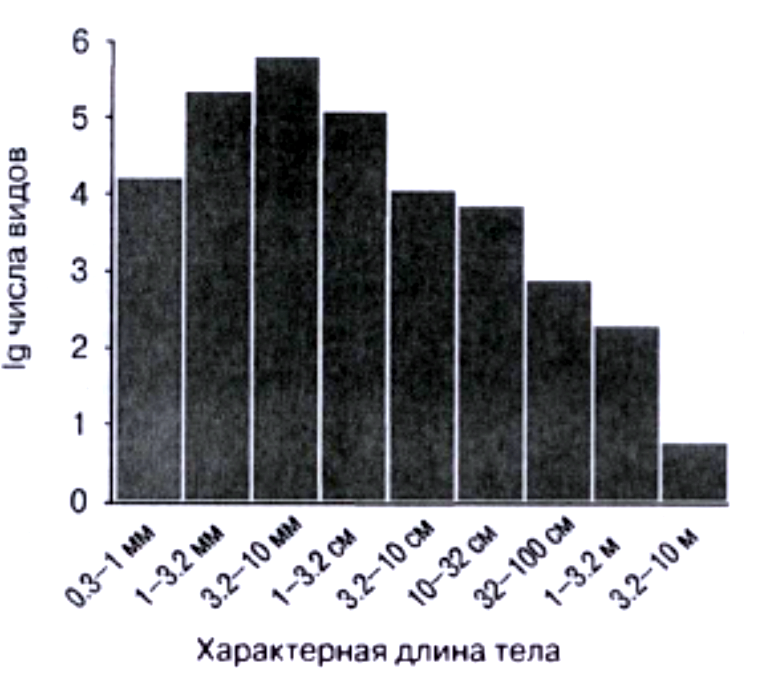
А.И. Азовский, опираясь на большой массив данных по видовому разнообразию организмов, обитающих на песчаной литорали Белого моря и на дне северных морей, выявил очень четкую закономерность: число видов, которое можно встретить на небольшом участке дна для мелких организмов гораздо выше, чем для крупных. Однако с увеличением площади исследования число мелких видов растет гораздо медленнее.
Данная зависимость — результат разного пространственного распределения мелких и крупных организмов. Мелкие виды характеризуются необычайно широким, порой космополитным распространением, и поэтому уже на относительно небольшой площади может обитать ощутимая часть мировой фауны (флоры). А вот крупные организмы нередко занимают относительно небольшие ареалы, совокупность которых удается охватить только при крупномасштабных обследованиях огромных акваторий.
Исследователи не раз задавались вопросом, чем объяснить необычайно широкое, порой всесветное распространение мелких организмов, прежде всего одноклеточных. Первые натуралисты, изучавшие пресные воды Новой Зеландии, были поражены, обнаружив там те же самые виды инфузорий и других простейших, которые обычны для водоемов Англии. Объясняется это следующими особенностями мелких организмов: небольшими размерами, очень высокой скоростью размножения, быстротой расселения, огромной численностью, малой вероятностью аллопатрического видообразования, т.е. возникновения в разных местах разных видов, происходящих от общего предка.
Мир, в котором живут существа микроскопических размеров, радикально отличается от мира, в котором живут существа более крупные. Размер тела, по которому проходит граница между двумя этими группами, может быть условно определен в 1 мм. Организмы более мелкие, как правило, характеризуются необычайно широким, зачастую космополитным распространением, а более крупные имеют ограниченные ареалы.
Видовое разнообразие организмов океана и суши¶
Хотя океан и занимает 2/3 поверхности земного шара, число видов организмов, обитающих в океане, существенно меньше, чем живущих на суше. Из каждых десяти известных науке видов девять — сухопутные. Систематики постоянно описывают новые виды, однако это соотношение не меняется. Причины различий в уровне биоразнообразия суши и океана в общем понятны.
Во-первых, хотя величины суммарной годовой первичной продукции океана и суши примерно равны, значительная часть акватории Мирового океана характеризуется крайне низкой продуктивностью и представляет собой, по сути, биологическую пустыню. Богатые видами сообщества животных могут сформироваться только при достаточно большом количестве пищи. Сообщество обитателей коралловых рифов, одно из самых богатых видами морских сообществ (на 1 га рифов может обитать до 300 видов коралловых полипов, 600 видов рыб и 200 видов водорослей), поддерживается высокой первичной продукцией самих кораллов, фотосинтезом водорослей, живущих непосредственно в клетках коралловых полипов. Прибрежные зоны океана характеризуются большим по сравнению с открытыми водами биоразнообразием и более высокой первичной продукцией.
Во-вторых, существенные различия в видовом разнообразии водных и наземных сообществ определяются особенностями самой среды обитания, ее физической структурой. Водная среда подвижна, она легко перемешивается, поэтому обитателям водной толщи, конкурирующим за общие ресурсы (например, элементы минерального питания в случае фитопланктона), разойтись по разным экологическим нишам достаточно сложно. Организмы не могут реализовать свое относительное преимущество, закрепившись в каком-либо месте. Для сообществ животных и растений, развивающихся на подводных поверхностях (организмы обрастания или обитатели дна), возможностей для дивергенции (расхождения признаков) гораздо больше, а дивергенция — необходимое условие видообразования.
На суше, как правило, растения гораздо лучше обеспечены элементами минерального питания (азотом и фосфором), что способствует развитию большой биомассы наземной растительности. В океане основные продуценты — организмы микроскопических размеров — цианобактерии и одноклеточные планктонные водоросли. Они быстро потребляются зоопланктоном, который в свою очередь служит основой всем последующим звеньям пищевой цепи.
На суше определяющая роль в развитии и поддержании необычайно высокого видового разнообразия многих сообществ (прежде всего сообществ насекомых и других мелких членистоногих) принадлежит наземной растительности. Суммарная поверхность стволов растений, ветвей и листьев — огромное пространство, в котором и существует множество мелких членистоногих. Более того, любое растение предоставляет мелким животным разные микроместообитания, пространственные ниши, по которым могут расходиться отдельные виды. Даже верхняя и нижняя стороны одного листа различаются условиями обитания.
Палеонтологи уже не раз отмечали, что резкий рост числа видов наземных организмов произошел примерно 110 млн лет назад, когда на суше началась быстрая эволюция покрытосеменных (цветковых) растений. Интенсивное видообразование происходило при этом не только среди самих растений, но и в других группах организмов, тесно с ними связанных, прежде всего грибов и многих животных. Среди последних — и опылители, и разносчики семян, и специализированные фитофаги, т.е. животные, питающиеся строго одним видом или, что чаще, несколькими близкими видами растений. Благодаря наличию специализированных опылителей (а их сейчас насчитывают около 200 тыс. видов) и распространителей семян стало возможным появление растений, которые не образуют крупных локальных скоплений, а разбросаны отдельными экземплярами по большой территории. Отсюда чрезвычайно высокое разнообразие деревьев даже на небольшом участке влажного тропического леса.
Оценка общего числа видов на Земле¶
Попытки оценить общее число видов, обитающих на нашей планете, предпринимались неоднократно. Цифры, полученные сведением данных, взятых из разных источников, всегда в той или иной степени приблизительны.
Во-первых, постоянно происходит описание новых видов, которые не успевают учесть в сводках.
Во-вторых, нередко один и тот же вид описывается разными авторами под разными названиями, и систематикам приходится время от времени пересматривать эти описания и выявлять синонимы.
Число видов в основных группах организмов
Грибы — 100 тыс. (скорее всего около 1 млн)
Сосудистые растения — 300 тыс.
Мхи — 16 тыс.
Кишечнополостные — 10 тыс.
Моллюски — 85 тыс.
Кольчатые черви — 16 тыс.
Паукообразные — 102 тыс.
Многоножки — 16 тыс.
Насекомые — около 1 млн Нематоды — 25 тыс.
Млекопитающие — 5,5 тыс.
Птицы — 10 тыс.
Пресмыкающиеся — 8,7 тыс.
Земноводные — 6,5 тыс.
Рыбы — 31,3 тыс.
Исследователи продолжают описывать новые виды, причем не только беспозвоночных. Так, за период, прошедший после 1950 года, новыми для науки признаны 297 видов птиц, 914 видов млекопитающих и 3418 видов земноводных. В последнем случае это более половины (55 %) от общего числа известных видов этой группы.
Конечно, больше всего описывают насекомых — около тысячи новых видов ежегодно. А поскольку их и так больше, чем всех остальных, вместе взятых, то понятны усилия исследователей, направленные на то, чтобы хотя бы примерно оценить общее число существующих сейчас видов насекомых (в том числе еще не описанных).
Чаще всего такие попытки основываются на определении числа видов, жизнь которых жестко связана с одним видом деревьев. Например, в начале 1980-х годов американский энтомолог Терри Эрвин в тропическом лесу в Центральной Америке (о. Барро-Колорадо, район Панамского канала) обработал ядовитым (содержащим инсектицид) дымом кроны нескольких деревьев одного вида и, собрав всех упавших членистоногих на растянутую внизу полиэтиленовую пленку, подсчитал общее число видов жуков, из которых многие, как он полагал, являются новыми для науки. Затем, приняв ряд допущений, он пришел к выводу, что строго специфическими, питающимися только на данном виде деревьев, можно считать 136 видов жуков (это самый богатый видами отряд насекомых), а всех членистоногих, которые могли быть связаны с одним видом деревьев, примерно 600. Поскольку общее число видов деревьев в тропических лесах — около 50 тыс., то перемножение этих цифр давало оценку в 30 млн видов.
Оценка Эрвина многими энтомологами была воспринята скептически. Ведь согласно его расчетам получалось, что большинство насекомых, находимых специалистами-систематиками в тропическом лесу, должны быть новыми для науки, хотя на самом деле это не так. Явно завышенной была и оценка числа видов насекомых, которые могут питаться только на одном виде деревьев.
Следующим этапом стала необычайно кропотливая работа, проделанная в дождевых лесах Новой Гвинеи чешским энтомологом Войтехом Новотным и его коллегами. В течение нескольких лет они обследовали деревья 51 вида и собрали с них около 50 тыс. растительноядных (точнее — листогрызущих) насекомых, относящихся к 935 видам. Преобладали жуки, гусеницы бабочек (чешуекрылых) и прямокрылые. Помимо простой констатации нахождения того или иного вида насекомых на листьях того или иного дерева, Новотный и его помощники оценивали полноценность корма в специальных экспериментах, выращивая разных насекомых на разных растениях.
В результате этой работы было показано, что подавляющее большинство насекомых в тропическом лесу питались несколькими видами растений. И если в среднем на одном виде растений кормилось около 30 видов жуков, 26 видов бабочек (гусениц) и 20 видов прямокрылых, то «узких специалистов», питающихся листвой только одного вида, среди них было существенно меньше: примерно 8 видов жуков, 13 видов бабочек и 3 вида прямокрылых. Таким образом, представление о чрезвычайной пищевой специализации тропических насекомых на поверку оказалось не более чем мифом. Расчетная оценка общего числа видов насекомых, полученная Новотным с учетом новых данных, это 4,9 млн, но никак не 30 млн. Согласно более поздним исследованиям общее число видов членистоногих в тропических лесах оценивается как 3,7 млн.
Ледниковые эпохи¶
Ледниковые эры в истории Земли¶
Интервалы холодного климата, в течение которых образуются обширные материковые ледниковые покровы и отложения длительностью в сотни миллионов лет, именуются ледниковыми эрами; в ледниковых эрах выделяются ледниковые периоды длительностью в десятки миллионов лет, которые, в свою очередь, состоят из ледниковых эпох — оледенений (гляциалов), чередующихся с межледниковьями (интергляциалами).
Геологические исследования доказали, что на Земле существовал периодический процесс изменения климата, охватывавший время от позднего протерозоя до настоящего времени.
В истории Земли выделяются следующие ледниковые эры:
- раннепротерозойская — 2,5–2 млрд лет назад;
- позднепротерозойская — 900–630 млн лет назад;
- палеозойская — 460–230 млн лет назад;
- кайнозойская — 30 млн лет назад–настоящее время.
Ледниковая эпоха оледенений (гляциалов) — отрезок времени геологической истории Земли, характеризующийся сильным похолоданием климата и развитием обширных материковых льдов не только в полярных, но и в умеренных широтах.
Особенности ледниковой эпохи:
- для неё характерны длительное, непрерывное и сильное похолодание климата, разрастание покровных ледников в полярных и умеренных широтах.
- ледниковые эпохи сопровождаются понижением уровня Мирового океана на 100 м и более, за счёт того, что вода накапливается в виде ледниковых покровов на суше.
- во время ледниковых эпох расширяются области, занятые многолетнемерзлыми породами, сдвигаются в сторону экватора почвенные и растительные зоны.
Установлено, что за последние 800 тысяч лет было восемь ледниковых эпох, каждая из которых продолжалась от 70 до 90 тысяч лет.
Кайнозойская ледниковая эра
Кайнозойская ледниковая эра (30 млн лет назад–настоящее время) — недавно начавшаяся ледниковая эра.
Настоящее время — голоцен, начавшийся ≈ 10000 лет назад, характеризуется как относительно тёплый промежуток после плейстоценового ледникового периода, часто квалифицируемый как межледниковье. Ледниковые щиты существуют в высоких широтах северного (Гренландия) и южного (Антарктида) полушарий; при этом в северном полушарии покровное оледенение Гренландии простирается на юг до 60° северной широты (т. е., до широты Санкт-Петербурга), фрагментов морского ледового покрова — до 46–43° северной широты (т. е. до широты Крыма), а вечной мерзлоты до 52–47° северной широты. В южном полушарии континентальная часть Антарктиды покрыта ледниковым щитом мощностью 2500–2800 м (до 4800 м в некоторых районах Восточной Антарктиды), при этом шельфовые ледники составляют ≈ 10 % от площади континента, возвышающейся над уровнем моря. В кайнозойской ледниковой эре наиболее сильным является плейстоценовый ледниковый период: понижение температуры привело к оледенению Северного Ледовитого океана и северных областей Атлантики и Тихого океана, при этом граница оледенения проходила на 1500–1700 км южнее современной.
Причины ледниковых эпох
Причины ледниковых эпох нераздельно связаны с более широкими проблемами глобальных климатических изменений, имевших место на протяжении истории Земли. Время от времени происходили значительные смены геологических и биологических обстановок. Следует иметь в виду, что начало всех великих оледенений определяется двумя важными факторами:
- во-первых, на протяжении тысячелетий в годовом ходе осадков должны доминировать обильные продолжительные снегопады.
- во-вторых, в районах с таким режимом осадков температуры должны быть настолько низкими, чтобы летнее снеготаяние сводилось к минимуму, а фирновые поля увеличивались из года в год до тех пор, пока не станут формироваться ледники. Обильная аккумуляция снега должна превалировать в балансе ледников на протяжении всей эпохи оледенения, так как если абляция превысит аккумуляцию, оледенение пойдёт на убыль.
Фирн — плотно слежавшийся, зернистый и частично перекристализованный, обычно многолетний снег. Абляция — уменьшение массы ледника или снежного покрова в результате таяния, испарения и механического удаления или разрушения.
Многолетняя мерзлота
Многолетняя мерзлота — часть криолитозоны, характеризующаяся отсутствием периодического протаивания. Общая площадь вечной мерзлоты на Земле — 35 млн км². Распространение — север Аляски, Канады, Европы, Азии, острова Северного Ледовитого океана, Антарктида. Районы многолетней мерзлоты — верхняя часть земной коры, температура которой долгое время (от 2–3 лет до тысячелетий) не поднимается выше 0 °C. В зоне многолетней мерзлоты грунтовые воды находятся в виде льда, её глубина иногда превышает 1000 м.
Вечная мерзлота — явление глобального масштаба, она занимает не менее 25 % площади всей суши земного шара. Материк, где вечная мерзлота отсутствует полностью, — Австралия, в Африке возможно её наличие только в высокогорных районах. Значительная часть современной вечной мерзлоты унаследована от последней ледниковой эпохи, и сейчас она медленно тает. Содержание льда в промерзлых породах варьируется от нескольких процентов до 90 %.
В Южном полушарии, за исключением Антарктиды, вечная мерзлота — элемент высокогорий. Наибольшая площадь (200 тыс. км2) — в Южной Америке, в Андах.
От 60 % до 65 % территории России — районы вечной мерзлоты. Наиболее широко она распространена в Восточной Сибири и Забайкалье.
Самый глубокий предел вечной мерзлоты отмечается в верховьях реки Вилюй в Якутии. Рекордная глубина залегания вечной мерзлоты — 1370 метров — зафиксирована в феврале 1982 года.
Уровень океана в ледниковую эпоху
Одно из самых ярких изменений в ледниковую эпоху — снижение уровня океана, вызванное не поднятиями суши, а уменьшением количества воды в океане. На Земле существует ограниченное количество воды. В любой момент времени часть её находится в море, часть — на пути к суше, в виде атмосферной влаги, а часть задерживается на суше в виде озёр и ледников или в виде грунтовых вод. Только некоторое количество воды возвращается обратно в море. Уровень моря в любой момент отражает соотношение между количеством воды, находящимся в каждом из этих четырёх состояний.
Когда в ледниковую эпоху температуры снижались, большая часть осадков выпадала в виде снега и меньшая — в виде дождя. Образовались ледники. Поскольку в море теперь возвращалось меньше воды, потому что большая часть её, участвующая в круговороте, накапливалась на суше в виде льда, глубина моря должна была уменьшаться.
Современный уровень моря определяется количеством воды, которое исключается из круговорота и существует в виде ледников Антарктиды и Гренландии, потому что где-либо в других местах льда совсем мало. Но 20000 лет назад образовавшиеся ледники распространялись в средних широтах так далеко, что на суше существовало огромное количество дополнительного льда. Рассчитав количество этого льда и зная площадь океана, мы можем определить, насколько должен был снижаться уровень моря в это время. Уровень океана должен был снижаться почти на 90 м.
Остатки растительности прибрежных лесов и болот, существовавших при более низком уровне моря, были обнаружены в буровых скважинах, заложенных в поисках нефти. Другие подобные остатки встречаются на поверхности морского дна; местами там находят пни и даже вертикально стоящие стволы деревьев с корневой системой. Рыболовными сетями с поверхности шельфа были извлечены зубы и кости животных ледниковой эпохи.
У тех участков побережья, где сейчас глубины малы, снижение уровня океана в ледниковую эпоху вызвало воссоединение островов с материком. Достоверно установлено, что это произошло с Британскими островами, которые оказались присоединены к Европе широкой равниной, смыкавшейся с Францией на юге и с Данией на востоке. По этой равнине в различное время свободно перемещались крупные млекопитающие — слоны, носороги, бегемоты, а также и более мелкие. На месте современного пролива Ла-Манш находилась долина реки, тёкшей к западу, а Рейн продолжался далее к северу, занимая долину, которая сейчас находится на дне Северного моря. Он впадал в океан значительно восточнее северной Англии.
Таким же образом Аляска соединялась с Сибирью, Сибирь — с Японией, Малаккский полуостров — с Суматрой, Явой, Борнео, а Австралия — с Новой Гвинеей и Тасманией. Хотя предположения об этом первоначально основывались только на существовании малых глубин в местах предполагаемых сухопутных мостов, но в большинстве случаев они были подтверждены находками одинаковых ископаемых животных по обе стороны «моста» — таких млекопитающих, которые могли попасть куда-либо только по суше.
Перигляциальная гиперзона
В плейстоцене наряду с покровными оледенениями существовала обширная перигляциальная гиперзона, которая распространялась на многие сотни, а иногда и тысячи километров от границы ледников и занимала значительную часть территории Евразии. Для перигляциальной гиперзоны был характерен очень суровый и засушливый климат с многолетней или мощной сезонной мерзлотой, скудной растительностью и арктической фауной, в которой формировались специфические формы рельефа и коррелятивные им осадки с характерными текстурами.
Наступление ледников, распространявшихся из Скандинавии, влекло за собой нарушение существующей зональности. Определяющим фактором этого процесса был не столько холод, сопровождавший ледник, сколько уменьшение влажности, проявлявшее себя не только в обрамлении ледника, но и на громадных расстояниях от него, вплоть до тропической зоны. На фоне иссушения климата лесная зона сокращалась и дробилась на части, которые продолжали уменьшаться в размерах. Открытые пространства расширялись за счёт уменьшения лесов. К максимальному развитию оледенения часть таких лесных «островов» исчезала полностью, а часть сохранялась в уменьшенном виде, образуя рефугиумы. На остальном огромном пространстве в обрамлении ледника устанавливалась перигляциальная гиперзона со своеобразными перигляциальными ландшафтами, которым нет аналогов в настоящее время: тундры, лесотундры, лесостепи, перигляциальные степи, тундролесостепи, редколесья.
Рефугиум — участок земной поверхности или Мирового океана, где вид или группа видов пережили или переживают неблагоприятный для них период геологического времени, в течение которого на больших пространствах эти формы жизни исчезли.
Тундростепь
Тундростепь — некогда обширная экосистема (биотоп) полярных и приполярных областей Евразии и Северной Америки эпохи плейстоцена. Участки реликтовых тундростепных ландшафтов — дриадовые тундростепи — сохранились до наших дней в Хакасии, на Алтае, отдельные представители этих растительных сообществ сохранились в Забайкалье и Прибайкалье.
Во время последнего ледникового максимума тундростепь — мамонтовая степь, была самым большим по площади биомом Земли. В длину она простиралась от современной Испании, на восток, через всю Евразию до Канады. Так же следует учитывать, что в тот период Евразия и Северная Америка были соединены посредством Берингии — сухопутного пути, на месте нынешнего Берингова пролива. В ширину — от современных арктических островов до Китая. Климат биома был холодный и сухой. В растительном покрове преобладали высококалорийные, кормовые сорта трав и кусты ивы. Они обеспечивали надёжную кормовую базы круглый год, для большого количества животных мегафауны. Основными травоядными того периода были — зубры, лошади, шерстистые мамонты.
Тундростепь имела стабильный, характерный климат и биосистему на протяжении 100 тыс. лет. Исчезла как биом 12 тыс. лет назад.
Тундростепь существовала по расширенным оценкам в датировке, между 126000–11700 лет назад. Последний ледниковый период достиг максимума между 33000–26500 лет назад. Затем в Северном полушарии началось постепенно потепление и отступление ледников, начиная с 19 тыс. лет назад, в Антарктиде — с 14500 лет назад.
Тундростепи находились на перигляциальных территориях с холодным и сухим климатом. Травяной ярус тундростепи формировали в основном не мхи (как в тундре), а злаки. Характерной чертой древних мамонтовых прерий было изобилие крупных животных: мамонты весом до 10 т, шерстистые носороги, лошади, бизоны и туры. Несмотря на сильные морозы зимой, за короткое жаркое лето в тундростепях успевало отрасти обильное разнотравье высотой до 2 м. Солнечная и сухая осень превращала его в «сено на корню», которым питались мамонты и другие травоядные животные во время долгой зимы.
Сухость климата являлась причиной низкого количества осадков и как следствие — малого количества облачных дней в году, в том числе и в летне-осенний период. Благодаря этому, осенью трава без риска отсыревания, высыхала под солнцем и ветром, дующим со стороны ледников. Так же, общее не высокое значение количества осадков, обеспечивало относительно малоснежные зимы, если сравнивать с нынешними временами. Малая глубина снежного покрова позволяла крупным животным мегафауны — мамонтам, шерстистым носорогам, лошадям, добывать корм, не тратя лишние силы на продвижение по глубокому снегу.
Нынешний климат в тундре и лесотундре намного суровее, к тому же, в силу большей влажности климата, осенью идут сильные дожди и поэтому травы сгнивает на корню, не превращаясь в сено. Так же, в силу большого количества осадков и облачных дней в году, произошло активное заболачивание огромных площадей. В этом причина того, что мхи вытеснили, как кормовая база, траву. Зимы так же в наше время намного суровее.
При обилии корма морозы им были не страшны, а от переохлаждения молодняк защищал толстый слой подкожного жира и густая лохматая шерсть. Животные мамонтовых степей являлись объектом охоты древних людей.
Главной чертой тундростепи было отсутствие лесов и почти полное отсутствие кустарников на огромных площадях. Сухой климат не позволял развиваться деревьям, таким как сосны, ели, берёзы, дубы. Когда климат начал постепенно теплеть и увлажнение возросло между 15000–11000 лет назад, степная растительность постепенно сменилась сначала кустарниками, затем между 12900–11700 лет назад — лиственными деревьями и затем тайгой.
При анализе остатков почвы тундростепей, которые сохранились в отложениях в вечной мерзлоте Сибири и Аляски, было установлено — содержание органических веществ, а значит плодородность почв тундростепи, были очень высокими, в сравнении с современным периодом в Сибири и других частях бывшей тундростепи. Почва давала базу для роста высококалорийных трав и развития большого количества животных.
По одной из версий, тундростепи исчезли из-за постепенного увлажнения и потепления климата, в результате которого ледник отступил на север, и произошли изменения в растительном составе тундростепей. Эти изменения привели к сокращению численности плейстоценовой мегафауны, и, в результате положительной обратной связи, стали необратимыми, что привело к зарастанию значительной части тундростепей кустарниками, и, впоследствии, хвойными лесами.
По другой, это произошло под воздействием человека, который сильно сократил популяцию основных видов-эдификаторов (мамонты, носороги и другие), которые сохраняли тундростепи, подъедая растения и вытаптывая поросль своими плоскими, массивными ступнями. После резкого сокращения их численности степь заменилась лесостепью, которую позднее сменила современная тундра.
Фауна плейстоцена в основном представлена современными, хорошо нам известными животными. Однако целый ряд видов в конце плейстоценовой эпохи исчез, что нередко считают результатом прямого истребления их древним человеком. Дело в том, что охотников привлекает прежде всего крупная добыча, а практически все вымершие животные относились к так называемой мегафауне, т.е. отличались большими размерами. Если к мегафауне условно отнести животных с массой тела во взрослом состоянии более 44 кг, то выясняется, что 50 тыс. лет назад таких было на Земле не менее 150 родов, однако 10 тыс. лет назад их было уже, по крайней мере, на 97 родов меньше. В Евразии и в Северной Америке вымерли мамонты, шерстистые носороги, дикие лошади (предки домашней лошади), пещерный медведь. В Северной и Южной Америке исчезли гигантские наземные ленивцы, а в Австралии дипротодон — самое крупное из когда-либо существовавших сумчатых. По габаритам и внешнему виду этот зверь напоминал гиппопотама, но на самом деле был родственником живущего и сейчас совсем небольшого вомбата. В наибольшей степени вымирание мегафауны коснулось Австралии, Южной Америки и Северной Америки. А вот в Африке оно было незначительным, что, по крайней мере отчасти, может быть объяснено тем, что Африка — родина человека. Его формирование как биологического вида происходило в тех экосистемах, элементами которых были и крупные животные. На других континентах человек был пришельцем, который по мере своего расселения по планете невольно вторгался в давно сложившиеся экосистемы.
В Северной Европе, Сибири и на Аляске вымирание мегафауны происходило в два этапа, определяемых крупными перестройками климата. Первая волна вымирания, наблюдавшаяся 45–20 тыс. лет назад, была связана с начавшимся сильным похолоданием. Тогда исчезли прежде всего теплолюбивые животные. Вторая волна была уже после Последнего Ледникового Максимума, 12–9 тыс. лет назад. Этот период характеризовался значительным потеплением, что в свою очередь привело к существенным изменениям ландшафта. На севере Евразии и на Аляске исчезли тундростепи (мамонтовые степи). Как говорит само название, мамонт был характерным видом фауны этих степей. Но кроме мамонтов бродили по мамонтовым степям и другие крупные травоядные млекопитающие; некоторые из них вымерли, например шерстистый носорог, но другие продолжают процветать и сейчас, например северный олень.
Для поддержания тундростепей была важна определенная климатическая обстановка. Хотя никто не сомневается, что первобытные охотники усиленно истребляли именно самых крупных представителей фауны, значение серьезных изменений климата никак нельзя сбрасывать со счетов. Во всяком случае на севере Евразии и Америки потепление, пришедшее на смену последнему оледенению, привело к резкому уменьшению территории тундростепей, на которых только и могли прокормиться мамонты, шерстистые носороги, дикие лошади и другие крупные травоядные. Открытые пространства быстро зарастали кустарниками, а затем и настоящим лесом. Очевидно, что общая численность популяций разных видов мегафауны сильно сократилась к тому времени, когда туда добрались, наконец, охотники.
Недавно это было убедительно показано на примере Аляски и Юкона. После Последнего Ледникового Максимума обширные территории там были заняты сухими мамонтовыми степями, на которых из крупных млекопитающих преобладали мамонты и лошади. Однако 13,5 тыс. лет назад, вслед за быстро произошедшим потеплением и увеличением количества выпадающих осадков, степи сменились зарослями кустарников. Появились бизоны и олени, в том числе лоси (а это уже лесные звери), лошади измельчали, а потом вымерли. Мамонты исчезли 12–11,5 тыс. лет назад. Возможно, что появившийся в это время человек сыграл определенную роль в исчезновении этого самого крупного представителя северной мегафауны, но если это и происходило, то на протяжении, по крайней мере, тысячи лет. Основной добычей древних охотников, судя по материалам археологических раскопок, были бизоны и олени.
Человек¶
Возникновение человека и его развитие, возникновение человеческого общества явилось результатом длительного развития живого вещества в пределах биосферы и стало существенным фактором перестройки конструкции земной биосферы.
Однако не следует преувеличивать воздействие человека на биосферу. Глобальные круговороты углерода, кислорода, азота и других важнейших для жизни элементов как определялись деятельностью бактерий, так и определяются ею сейчас. К бактериям как важнейшим двигателям круговоротов прибавились позже (примерно 400 млн лет назад) только наземные растения и грибы. Деятельность человека, конечно, влияет на глобальные круговороты углерода, азота и фосфора, но, как правило, результаты этого влияния сводятся только к некоторым модификациям. Основное воздействие человека на биосферу проявляется в другом — прежде всего в уничтожении сложившихся природных ландшафтов и замене их ландшафтами, окультуренными — полями, плантациями, пастбищами. Если не учитывать территории, покрытые ледниками, то под влиянием человека оказывается примерно 50 % всей оставшейся поверхности суши. Производство пищи по своим масштабам — уже биосферный процесс.
Эволюция человека¶
Огромный прогресс в изучении ископаемых предков человека связан как с внедрением новых методов исследования, что привело к расшифровке геномов ныне живущих и некоторых вымерших гоминид, так и с расширением работ по поиску новых ископаемых останков древних людей и их родственников.
Из ныне живущих человекообразных обезьян ближайший наш родственник — шимпанзе. Самих шимпанзе разделяют сейчас на два вида: обыкновенный шимпанзе и карликовый шимпанзе, или бонобо. Но линии этих двух видов разошлись сравнительно недавно — 1,5–2 млн лет назад, а вот линии сапиенсов и шимпанзе разделились гораздо раньше, примерно 6–7 млн лет назад. Это разделение совпало по времени с довольно резким изменением климата в Восточной Африке — похолоданием и аридизацией. Леса стали отступать (деревьям всегда нужно много влаги), а их место заняла саванна, где открытые участки, занятые травянистой растительностью, чередовались с разреженным лесом. Один из разошедшихся видов (шимпанзе) так и остался лесным обитателем, как его предки, а вот другой, давший линию, приведшую в конце концов к сапиенсам, вышел на открытую местность. При этом ему пришлось встать на две ноги и выпрямиться во весь рост.
До недавнего времени самым далеким достаточно хорошо изученным двуногим предком человека считалась знаменитая Люси, жившая в Восточной Африке 4,0–3,9 млн лет назад и отнесенная к виду Australopithecus afarensis. Но в 2009 году вышел специальный номер журнала Science, посвященный находке скелетных останков самки гоминид, получившей имя Арди, — представителя более древнего вида Ardipithecus ramidus, жившего 4,4 млн лет назад. Хотя Арди передвигалась в основном на двух ногах, обитала она все же в лесу (возможно, разреженном), а не в открытой саванне.


Переход к двуногому хождению, был очень важным шагом в становлении древнего человека:
- во-первых, это давало возможность видеть окрестности, что в высшей степени актуально для любого млекопитающего, живущего на открытой местности. Только так можно было найти достаточно крупную добычу и вместе с тем вовремя заметить тех хищников, которые сами представляют большую опасность и от которых надо держаться подальше.
- во-вторых, на двух ногах можно было гораздо быстрее бегать — ведь предкам человека, обитавшим в лесу, нужно было прежде всего хорошо лазать по деревьям, а не бегать.
- в-третьих, передние конечности (руки) оказывались свободными и ими легко можно было взять палку, камень, а потом научиться совершать и куда более сложные действия.
Несколько миллионов лет назад разных видов двуногих гоминид, которых условно уже можно назвать людьми, существовало довольно много, а судя по тому, как быстро находят все новых и новых представителей, нам известна только очень небольшая их часть. Почти два миллиона лет продолжалось существование на Земле одного из наших предков — Homo erectus, человека прямоходящего. Эректус оказался весьма успешным видом, так как достиг высокой численности и из Африки широко распространился по Евразии, дойдя до территории современного Восточного Китая (останки эректусов там известны как синантропы). Эректусы изготавливали каменные орудия, сначала довольно примитивные, а затем и куда более совершенные. Они, по-видимому, пользовались огнем, хотя вряд ли могли его добывать сами.
Последние эректусы существовали тогда, когда уже появились первые сапиенсы и неандертальцы. Но до этого от Homo erectus чуть менее миллиона лет назад должна была отделиться линия, приведшая к возникновению гейдельбергского человека (Homo heidelbergensis). О существовании гейдельбергского человека мы судим в основном по многочисленным оставшимся каменным орудиям, костных же фрагментов сохранилось мало. Примерно 400–300 тыс. лет назад от гейдельбергского человека началась линия неандертальцев (Homo neanderthalensis). Возможно, это имело место в Передней Азии, после чего неандертальцы заселили Европу и распространились далеко на восток вплоть до юга Сибири. В Европе они существовали еще 40–30 тыс. лет назад и вымерли, по-видимому не выдержав конкуренции с сапиенсами. Будучи очень крепкого сложения, неандертальцы легче, чем другие гоминиды, приспосабливались к более холодному климату.
Другая линия гейдельбергских людей, отделившаяся 300–200 тыс. лет назад, впоследствии дала сапиенсов. Самые ранние хорошо сохранившиеся костные останки настоящих сапиенсов найдены на юге Эфиопии и датированы весьма точно: 195 тыс. лет назад.
Совершенствование методов молекулярной генетики позволило расшифровать (хотя и не полностью) геном неандертальца — нашего ближайшего родственника среди вымерших гоминид. Сопоставление его с геномами современных людей показало, что вскоре после разъединения этих видов они все же изредка скрещивались. По подсчетам специалистов, неандертальских генов у нас 1–4 %. Но присутствуют они только у европейцев и азиатов. А вот у коренных обитателей Африки их нет. Данное обстоятельство — свидетельство того, что встреча сапиенсов и неандертальцев произошла не в Африке, а, видимо, где-то в Передней Азии уже после того, как некоторая группа сапиенсов мигрировала из Африки на восток, начав свое постепенное расселение по Евразии.
Первые сапиенсы, обитавшие в Восточной Африке, довольно быстро увеличивали свою численность, что не могло не приводить к напряженной внутривидовой конкуренции за места обитания (прежде всего — территорию, где можно добыть пищу). Кроме того, конкуренция, видимо, была и с другими жившими там гоминидами. Во всяком случае, примерно 125 тыс. лет назад сапиенсы мигрировали на север, через тогда еще зеленую Сахару до устья Нила, а затем на восток, в близлежащие районы Леванта. Эта популяция позднее, примерно 90 тыс. лет назад, вымерла при очередном сильном похолодании. Второй «исход» сапиенсов из Африки, примерно 85 тыс. лет назад, был более удачным, хотя начало его было довольно сложным: Красное море обходили не с севера, как в первый раз, а с юга. При этом нужно было переправиться через Баб-эль-Мандебский пролив, соединяющий Красное море с Аденским заливом Индийского океана. Сейчас этот пролив — глубокий и отличающийся быстрым течением, но были периоды в недавней истории Земли, когда из-за падения уровня океана он превращался в череду отмелей. Правда, исследователи отмечают, что сильное понижение уровня океана произошло в период максимума оледенения, а это было уже значительно позже. Так что загадка остается.
Переправившись на юг Аравийского полуострова, древние люди начали свой долгий путь на восток по берегу Индийского океана. Затем они обогнули полуостров Индостан, далее остров Борнео, который тогда был еще не островом, а соединялся с материком, и, так же двигаясь по берегу уже Тихого океана, достигли современного Восточного Китая. Расселение сапиенсов по Евразии было прервано природной катастрофой — чудовищным по силе извержением вулкана Тоба на Суматре, произошедшим 73000 лет назад. Объем выброшенного тогда материала более чем в 100 раз превышал выбросы вулкана Тамбора в Индонезии при извержении 1815 года, крупнейшем известным для исторического времени, приведшем к очень сильному похолоданию в Северном полушарии. В результате извержения вулкана Тоба значительная часть Южной Азии покрылась слоем вулканического пепла толщиной 15 см. Средняя температура для всей поверхности Земли понизилась на 3–5 °С, что подтверждается и данными анализа ледовых кернов из Гренландии. Вся жизнь на огромной территории Юго-Восточной Азии была уничтожена. Численность сапиенсов, обитавших за пределами Африки, резко снизилась.
По-видимому, именно тогда и наблюдалось прохождение сапиенсов через «бутылочное горлышко» низкой численности, когда вся популяция сапиенсов насчитывала от 3 до 10 тыс. человек. Вот от этой популяции, жившей, возможно, где-то в Передней Азии 60 тыс. лет назад, пошло новое расселение человека по берегу Индийского океана вплоть до Австралии. Естественно, что такая небольшая группа особей отличалась малым генетическим разнообразием в сравнении с той популяцией, возможно, даже несколькими популяциями сапиенсов, которые продолжали жить в Африке. Более того, по мере расселения людей по Евразии их генетическое разнообразие снижалось все в большей и большей степени, что сейчас доказано многочисленными данными молекулярной генетики, а также анализом измерений черепов. Причина такого снижения разнообразия понятна — ведь при своем продвижении на восток не вся популяция снималась с места и мигрировала. Всегда это была какая-то часть, выборка, из которой при продолжении миграции была новая выборка — и так далее.
В Австралию люди проникли 45 тыс. лет назад. Значительная часть Евразии заселялась в период от 45 до 25 тыс. лет назад. Переселение людей из Азии в Америку происходило по сухопутному мосту — Берингии, соединявшей Чукотку и Аляску. Началось оно, скорее всего, в промежутке между 22 и 16 тыс. лет назад, а 15 тыс. лет назад люди присутствовали уже как в Северной, так и в Южной Америке. Таким образом, к этому времени Homo sapiens обитал на всех континентах. Ни один другой представитель гоминид не смог достичь такого успеха.
Каковы причины столь стремительного расселения Homo sapiens по нашей планете? Почему ни один из его предшественников, в том числе эректус и неандерталец, сумевшие значительно увеличить ареал своего первоначального распространения, не смогли достичь подобного успеха? Прежде чем пытаться ответить на этот вопрос, надо понять, что заставляло определенную часть популяции сниматься с места и двигаться в еще неизведанные края. Здесь исследователи в общем единодушно говорят о двух причинах:
- истощение ресурсов в местах большого скопления людей. Не следует забывать, что все они были охотниками-собирателями, а для того, чтобы прокормить семью (в которой на самом деле все, кроме малых детей, участвовали в добывании пищи), им требовалась довольно обширная территория. Пищевых ресурсов на этой территории постепенно становилось все меньше и меньше, что заставляло человека двигаться в новые места.
- заболевания, неизбежно возникавшие в местах скопления большого числа людей. Патогенные организмы долгое время сохранялись в жилищах и около них. Недавнее эпидемиологическое исследование, проведенное во временных поселениях пигмеев в Центральной Африке, показало, что пробы почвы, взятые с утоптанных и тщательно выметенных площадок перед хижинами, содержат очень много патогенных микроорганизмов. Древние люди, конечно, не могли знать о механизмах возникновения заболеваний, но некий эмпирический опыт накапливался. Кроме того, работал и демографический фактор: в популяциях, долго проживающих на одном и том же месте, рождаемость снижалась, а смертность порой была даже выше, чем в группе мигрантов.
Истощение ресурсов и рост заболеваемости — факторы, которые действовали по отношению не только к сапиенсам, но и к другим расселяющимся гоминидам. Успех же человека, очевидно, объяснялся некими особенностями, которые не были столь хорошо выражены у его ближайших родственников. Речь идет прежде всего о необычайно быстром и успешном развитии «культуры», под которой антропологи подразумевают передачу из поколения в поколение знаний и умений не генетическим, врожденным путем, а через обучение, путем наблюдения за соседями и родственниками. Важно и то, что родители, носители определенных культурных навыков, специально передавали свой опыт детям. В зачаточной форме культура есть и у ныне живущих обезьян. В частности, много культурных навыков, причем нередко разных в отдельно живущих группах особей, выявлено у шимпанзе. Эти обезьяны показывают своим детям, как использовать камни для разбивания твердой скорлупы орехов или как с помощью очищенной палочки можно доставать вкусных термитов, как строить гнезда на деревьях для ночевки и так далее.
Примерно 45–35 тыс. лет назад произошло то, что археологи традиционно называли «верхнепалеолитической революцией», — скачок в культурном и технологическом развитии человека. Именно тогда сапиенсы проникли в Европу и вытеснили неандертальцев. Но скачок был не таким резким, а весьма совершенные орудия человек начал изготавливать даже раньше. Однако по мере расселения его по Евразии, а потом и по Америке каменные орудия становятся особенно хорошо обработанными, более изощренными и приспособленными для строго определенных целей. Чтобы изготовить такое орудие, нужно не только перенимать опыт соплеменников, но и самому в каждом случае проявлять немалую сообразительность и умение координировать сложные манипуляции.
Безусловно, немаловажное значение имело и то, что свою первую длительную миграцию за пределы родной Африки человек совершал по побережью Индийского океана. Во-первых, по берегу передвигаться значительно легче. Во-вторых, на берегу, в литоральной (приливно-отливной) зоне, всегда можно найти пищу, прежде всего моллюсков. Кучи оставшихся раковин маркируют путь древнего человека.
Когда древние люди оказались не в густом тропическом лесу, а в саванне, где участки леса перемежались с открытыми участками, они к собирательству прибавили охоту. В это время саванна была населена множеством крупных животных, как травоядных, так и хищников. Слоны, бегемоты, носороги были, конечно, слишком крупной добычей для человека (хотя потом он научился расправляться и с ними). Бродили по саванне и различные копытные: полорогие (всевозможные антилопы), а также лошади (зебры). Кроме того, были крупные свиньи. И очень много хищников, преимущественно представителей семейства кошачьих, а также гиеновых. Большинство этих хищников уже давно вымерли, хотя некоторые остались, например, лев. Идущие на охоту люди всегда должны были опасаться, как бы самим не стать добычей хищников. Охота была возможна только при высокой степени согласованности действий и использовании достаточно совершенных орудий.
Но, по-видимому, основным способом получения мясной пищи древними людьми была все же не настоящая охота, а использование свежей падали, остатков пиршества хищников. Особую «услугу» человеку оказывали саблезубые кошки. Дело в том, что у саблезубых кошек очень своеобразное строение зубного аппарата. Он великолепно приспособлен для умерщвления добычи, разрывания кожи и отрывания крупных кусков мяса. Но вот обгладывать кости саблезубые кошки не могли. Поэтому оставшиеся недоеденные туши антилоп, зебр и свиней служили лакомой добычей для падальщиков. Особенно эффективным хищником был мегантереон — саблезубая кошка с массой тела около 100 кг. Более крупная, весящая около 190 кг саблезубая кошка, гомотерий, представляла серьезную опасность для древнего человека. Нашим предкам приходилось быть очень осторожными, наблюдая за завалившими антилопу саблезубыми кошками, чтобы потом забрать свою порцию мяса и уйти. Это, конечно, возможно только при согласованных действиях коллектива охотников и наличии хороших инструментов для быстрой разделки туши. Куски мяса нужно было сразу же унести и спрятать в надежном месте. Важно и то, что основные конкуренты человека в потреблении падали, гиены, вели преимущественно ночной образ жизни, а человек был дневным существом. Кроме того, гиены плохо бегали, и древние люди вполне могли от них убежать.
Значительное увеличение содержания мяса в рационе древнего человека привело к тому, что его пища стала более калорийной, при этом на сам процесс добывания пищи стало тратиться меньше времени. Соответственно, высвободилось время на другую важную активность, например, изготовление все более совершенных каменных орудий. Все это способствовало развитию мозговой деятельности и лучшей коммуникации между членами совместно живущей группы. Особенно важным было постепенное введение практики использования огня.
Существует довольно много примеров того, что огнем пользовались уже эректусы. Но первый настоящий очаг, найденный на территории современного Израиля, который датируется довольно точно, имеет возраст 790 тыс. лет. Очевидно, древние люди (еще не сапиенсы, но их непосредственные предки — гейдельбергские люди) умели поддерживать огонь в этом очаге и пользовались им многократно. Приготовленное на огне мясо усваивалось гораздо лучше, что в свою очередь положительно сказывалось на общем качестве питания. Овладение огнем — важнейший шаг как в развитии самого человека, так и в расширении его воздействия на окружающую природу.
Будучи охотником-собирателем, владеющий огнем и умеющий делать все более совершенные каменные орудия Homo sapiens постепенно осваивал все континенты. Переход к оседлому образу жизни происходил постепенно, но уже 11 тыс. лет назад, а возможно, и раньше, 13 тыс. лет назад, началось одомашнивание и введение в культуру нескольких видов растений. Конечно, сначала люди просто собирали семена дикорастущих растений (например, ближайших родственников ячменя, пшеницы и ржи) и перетирали их на камнях в муку. Жернова, ступка и пестик возникли раньше, чем какие-то растения были введены в культуру. Но постепенно происходил отбор по тем признакам, которые представлялись человеку наиболее ценными.
По-видимому, введение в культуру злаковых хлебных культур произошло впервые 13–10 тыс. лет назад в пределах региона, известного среди археологов как Плодородный полумесяц. На карте он выглядит действительно как полумесяц, охватывающий низовья Нила, земли Леванта (территория на востоке Средиземноморья, где сейчас располагаются Сирия, Ливан, Израиль, Иордания, Турция) и Месопотамии. На этой территории весной выпадает довольно много осадков (относительно окружающих ее очень засушливых регионов), а почвы характеризуются высоким плодородием. Плодородный полумесяц связывает районы Древнего Египта и Месопотамии, и поэтому его справедливо считают колыбелью человеческой цивилизации.
Первыми растениями, введенными в культуру в разных местах Плодородного полумесяца, были злаки: ячмень, беотийская пшеница, пшеница двузернянка и рожь, а также бобовые: чечевица и горох. Поскольку переход от собирания семян дикорастущих растений до введения их в культуру может быть весьма растянут во времени, датировать по археологическим данным, когда именно произошло одомашнивание, довольно трудно. Недавно, правда, были получены данные при новых раскопках, проведенных немецкими и иранскими археологами на территории Западного Ирана, в горах Загроса. Авторам этой работы повезло: в помещении, которое они раскопали, издавна обрабатывали собранные семена, причем время от времени там делали пол из глины. Соответственно, отдельные археологические слои оказались четко разделенными. В наиболее древних слоях (примерно 11 тыс. лет назад) довольно много остатков дикого ячменя. Позднее, примерно 9800 лет назад, все в большем количестве начинают встречаться колоски окультуренной пшеницы двузернянки — крупные ее семена уже не могли распространяться сами, без помощи человека. В западных районах Плодородного полумесяца это произошло несколько раньше, 10500–10000 лет назад. Примерно тогда же в этих районах одомашнили козу.
Область Плодородного полумесяца была первая, но не единственная, где начали вводить в культуру местные растения. Н. И. Вавилов выделял семь территорий (порой довольно обширных), где происходило введение в культуру тех или иных растений. Помимо области Плодородного полумесяца, это Мезоамерика (центральная и южная часть Мексики и прилегающие части Центральной Америки), давшая кукурузу (9–8 тыс. лет назад), а до этого еще тыкву, восточная часть Северной Америки, где в культуру был введен подсолнечник (5 тыс. лет назад), Южная Америка, где в разных местах одомашнили картофель, батат и арахис. В Восточном Китае были окультурены рис и просо. В Африке южнее Сахары — сорго и африканский рис. В высокогорной части Новой Гвинеи — банан.
Область Плодородного полумесяца, безусловно, имела особое значение как родина основных зерновых культур, распространившихся по всей Евразии. И дело не только в благоприятном средиземноморском климате, но и в составе растений. Обилие однолетних злаков, часто произрастающих большими куртинами и дающих большое количество достаточно крупных семян, позволяли вполне успешно развиваться примитивному сельскому хозяйству — исключительно за счет собирания и обработки семян естественно растущих, еще не культивируемых специально растений. Это было первым шагом к их последующему культивированию.
Сельскохозяйственные культуры из центров своего происхождения постепенно распространялись по всем континентам, за исключением Австралии. Примерно 2500 лет назад на большей части территорий, где обитал человек, уже существовало сельское хозяйство. Так, пшеница двузернянка из области Плодородного полумесяца, введенная там в культуру 10,5 тыс. лет назад, уже через 2 тыс. лет достигла территории Греции, а еще через тысячу лет — территории современной Германии.
Параллельно с введением в культуру определенных растений происходило и одомашнивание животных. Видимо, первыми домашними животными стали: коза, ее диким предком был безоаровый козел, и овца, ее диким предком был азиатский муфлон. Оба эти вида сохранились в природе и до сих пор широко распространены в Юго-Западной Азии. Видимо, несколько позднее и в разных местах независимо произошло одомашнивание тура, давшего крупный рогатый скот (корову). Тур был широко распространен в Евразии. Его ареал охватывал Европу и тянулся широкой полосой на восток вплоть до Тихого океана. Особый подвид тура обитал в Северной Африке. Последние дикие туры еще 3 встречались в Европе совсем недавно, в XVI веке. В долине Инда появился свой рогатый скот — зебу, который называют еще «горбатым скотом», поскольку происходит он от дикого зебу, давно уже отделившегося от тура. В Китае была одомашнена свинья (предок — дикий кабан). Затем, скорее всего, независимо кабан был одомашнен и в Европе. В Мезоамерике одомашнили только индейку. Подходящих для одомашнивания млекопитающих в Америке просто не оказалось (только позднее в Южной Америке были одомашнены лама, альпака и морская свинка).
Первоначально всех домашних животных использовали только на мясо (не считая шкуры и кожи, которые шли на одежду). Но вскоре коз, овец и крупный рогатый скот стали доить. Полученное молоко жившие тогда люди во взрослом состоянии усваивать не могли. Фермент лактаза, расщепляющий содержащийся в молоке сахар, лактозу, образовывался у них только в раннем детстве, в период грудного вскармливания. У взрослых людей лактаза не вырабатывалась и молоко не усваивалось — употребление его в пищу приводило к серьезным заболеваниям желудочно-кишечного тракта. Только значительно позже у некоторых народов, где регулярно использовали разные обработанные молочные продукты (например, на севере Европы), постепенно произошли генетические изменения, позволившие людям и во взрослом состоянии пить молоко. У них лактаза не прекращала вырабатываться в течение всей жизни. Это классический пример быстро протекавшей эволюции самого человека. Но при этом в любой популяции остается некоторая часть (5–10 %) людей, неспособных усваивать молоко после периода младенчества. У тех же народов, где молочные продукты не использовали, эта доля составляет 90 % и даже 100 % (у многих южных народов или у индейцев Северной Америки).
Трудности усвоения молока можно преодолеть, если употреблять в пищу не непосредственно молоко, а полученные из него сбраживанием продукты типа йогурта, а также сыра, в которых содержание лактозы резко снижено (ее использовали молочнокислые бактерии). Неудивительно, что среди керамики, происходящей из районов с развитой культурой молочного хозяйства, столь часто встречается посуда с дырочками: она как раз удобна для производства сыра. Недавно убедительные свидетельства использования продуктов, полученных из молока крупного рогатого скота, обнаружены в Сахаре. Замечательные наскальные рисунки, изображающие диких животных, коров, а рядом с ними людей, были известны уже давно, но только недавно тщательное обследование современными методами аналитической химии выявило на стенках найденной там же керамической посуды следы специфических жиров из молока. Надо, конечно, напомнить, что в то время, примерно 7 тыс. лет назад, Сахара была еще зеленой, т.е. вовсе не пустыней, а саванной с присущей ей довольно богатой растительностью и животным миром.
К сожалению, одомашненные животные стали источником не только ценных продуктов питания, но и ряда болезней. При оседлом образе жизни люди невольно тесно контактировали с домашними животными и их выделениями, а соответственно с микроорганизмами, являющимися возбудителями тех или иных заболеваний. Конечно, при переходе в организм человека патогенные микроорганизмы эволюционировали, приспосабливаясь к новой для них среде. Методы современной молекулярной генетики позволяют установить эти связи. Так, возбудитель чумы крупного рогатого скота эволюционировал, став возбудителем кори у человека. Оспа, видимо все же в ослабленной форме, также перешла к человеку от коров (у которых есть своя, коровья оспа) или какого-то другого скота. Грипп (в Китае) получен от свиней и уток, коклюш — от свиней и собак.
Процесс получения людьми от животных возбудителей, превратившихся в смертельно опасных для человека, но не являющихся таковыми для их исходных хозяев, продолжался еще долгое время. К примеру, печально известный ВИЧ, вирус, вызывающий очень тяжелое заболевание, в крайней форме переходящее в СПИД, уже во второй половине XX века перешел к человеку от обезьян, добытых охотниками в качестве источника пищи.
Но первые возбудители, полученные древними людьми, перешедшими к оседлому образу жизни, были не столь опасны: смертность была, но она не косила всех подряд — у значительной части населения вырабатывался иммунитет. А вот охотники-собиратели, приходившие в поселения земледельцев и скотоводов, оказывались совершенно неподготовленными к новым для них болезням и часто погибали. Отношения между охотниками и теми, кто жил в поселениях, порой были весьма напряженными, хотя в начале развития земледелия члены племен охотников-собирателей просто время от времени возвращались к своим посевам, пропалывали их, собирали урожай, обрабатывали его и пытались как-то сохранить, а потом снова уходили в горы, где занимались охотой и собирательством.
Мы бы теперь сказали, что охотники-собиратели вели более здоровый образ жизни. Наоборот, перешедшие к оседлому образу жизни постоянно существовали в условиях очень высокой скученности. Смертность была высокой, особенно среди детей и подростков. Основная причина — заболевания, получаемые как от контактов с животными, так и друг от друга. Рождаемость была, однако, также весьма высокой: промежуток между рождениями детей составлял всего два года. Постепенно оседлый образ жизни распространялся по всему миру. Особенно быстро это происходило в Евразии. Там переход к оседлости, возможный только при достаточно развитом сельском хозяйстве, возникал независимо в разных местах. Это всегда было перемещение людей вместе с введенными в культуру растениями и одомашненными животными. Распространение людей — всегда и распространение определенных знаний и умений, всего того, что мы называем культурой.
Единственным континентом, не затронутым ни земледелием, ни скотоводством, оставалась Австралия, хотя она и была заселена людьми очень давно. Обитатели Австралии продолжали вести образ жизни охотников-собирателей. Правда, они научились использовать огонь, вызывая специальные поджоги растительности, после которых проростки съедобных растений начинали пробиваться из-под земли.
Неолитической революцией археологи обычно называют переход от охоты и собирательства к сельскому хозяйству. Границы этого периода во времени четко не определены, поскольку в разных местах земного шара указанный переход начался в разное время (от 12 до 3 тыс. лет назад) и шел с разной скоростью. Твердо доказанным пока можно считать, что зародилось земледелие и скотоводство около 10–12 тыс. лет назад на Ближнем Востоке, в пределах Плодородного полумесяца. Окончанием неолитической революции иногда условно считают появление первых шумерских городов (Месопотамия) 6–4 тыс. лет назад. Шумерская цивилизация, будучи исторически первой цивилизацией на Земле, отличалась уже необычайно высоким уровнем культуры, в частности появлением письменности и ряда технологических достижений, радикально усиливших воздействие человека на природу. Прежде всего это изобретение колеса, плуга (скот использовали как тягловую силу) и весьма совершенная система ирригации.
Неолитическая революция ознаменовалась демографическим сдвигом: при оседлом образе жизни возрастала не только рождаемость, но одновременно и смертность, особенно в возрастной группе от 5 до 19 лет. Численность таких популяций увеличивалась очень медленно, поскольку высокая рождаемость сводилась почти на нет высокой смертностью. Тем не менее расселение земледельцев и культивируемых ими растений продолжалось. Собирательство и охота могли поддержать существование не более 0,05 человек на 1 км2. Для сравнения укажем, что современное сельское хозяйство может с 1 км2 возделанной земли обеспечить пропитание 54 человек.
Общую численность людей на Земле на заре развития сельского хозяйства специалисты оценивают в 6 миллионов. В 2011 году она достигла 7 миллиардов, увеличившись, таким образом, за 11 тыс. лет в 1200 раз. Рост народонаселения в течение всего этого периода был очень неравномерным. В доиндустриальную эру он составлял всего 0,1–0,2 % в год, поскольку рождаемость почти уравновешивалась смертностью. В XIV веке в Европе разразилась чума («черная смерть»), унесшая жизни, по разным оценкам, от 75 до 200 млн человек. Чума пришла из Китая через Великий шелковый путь. Чуть более быстрый рост народонаселения отмечен в начале XVIII века, но стремительный взлет начался только в 1950-х годов за счет радикального снижения детской смертности в развивающихся странах.
Максимальную скорость относительного прироста народонаселения за всю историю человечества (2,1 % в год) наблюдали в 1960-х годах. Затем скорость прироста стала снижаться (в 2000 году уже только 1,35 % в год). Точка перелома кривой роста была пройдена, но общая численность народонаселения продолжала (и сейчас продолжает) расти.
Серьезные сдвиги произошли за последние два столетия по части вклада разных стран в общий прирост населения Земли. Если в XIX веке он происходил в основном за счет европейских стран и США, то в середине XX решающее значение стали иметь развивающиеся страны Центральной Америки, Юго-Восточной Азии и Африки. Именно в этих странах произошел демографическии переход, который и обеспечил необычайно быстрый рост популяций. Суть перехода в том, что в течение долгого времени в этих странах высокая рождаемость практически полностью нивелировалась столь же высокой смертностью, младенческой и детской. Разность рождаемости и смертности была столь незначительна, что численность населения росла крайне медленно. Но в 1960-х годах в развивающихся Странах (при поддержке развитых стран) удалось поднять уровень здравоохранения, в результате чего младенческая и детская смертность существенно снизились, однако рождаемость некоторое время оставалась еще очень высокой, так что скорость роста населения резко возросла.
О рождаемости в демографии судят по коэффициенту фертильности. Согласно более точному развернутому определению коэффициент фертильности — среднее количество детей, которое родила бы женщина гипотетического поколения за всю жизнь при условии сохранения в популяции того же распределения рождаемости по возрастам, которое наблюдалось в данном регионе в данный год.
За последние 50 лет фертильность в целом по миру (в том числе в Китае и Индии, где живет наибольшее количество людей) заметно снизилась, но разные страны очень сильно различаются по этому показателю. Высокий уровень фертильности — в среднем более 5 детей на одну женщину — сохраняется в странах Африки, расположенных к югу от Сахары. Так, в Анголе он составляет 5,3, а в Республике Нигер — 7,5. В азиатских странах фертильность ниже (2,6 в Индии, 1,6 в Китае, 1,4 в Японии, но 6,2 в Афганистане). В странах Европы и в России фертильность очень низкая — от 1,1 до 2,2. При сохранении низкой смертности для поддержания численности популяции на том же самом уровне необходимо, чтобы значение фертильности было по крайней мере 2,1–2,2.
Именно от того, какой будет средняя фертильность, зависит прогноз дальнейшей динамики народонаселения Земли. Если она будет сохраняться на современном уровне, то общая численность народонаселения достигнет 9,3 млрд к 2050 году и 10,1 млрд к 2100 году. Если допустить, что фертильность будет снижаться, то в 2050 году ожидается только 8,5 млрд Разные модели в зависимости от принятых значений будущей фертильности дают прогноз на 2100 год от 6,1 до 15,8 млрд Если говорить о максимальной численности народонаселения, по достижении которой дальнейшего роста уже не происходит, то при низкой фертильности это произойдет уже в 2045 году (максимальное значение — 8,1 млрд), а при высокой — после 2100 года (15,8 млрд).
Разные страны значительно различаются по уровню уже достигнутой численности народонаселения. Наибольшее количество людей проживает в Китае (1,35 млрд). От него только слегка отстает Индия (1,24 млрд), которая к 2050 году станет самой многолюдной страной — с общей численностью народонаселения 1,69 млрд Из 10 самых крупных по численности населения стран только три — США, Россия и Япония — относятся к категории развитых. Но вскоре Россия уже не будет относиться к первой десятке самых населенных стран, а вот Нигерия, которая сейчас занимает седьмое место, перейдет на четвертое.
Очень важную информацию, позволяющую судить о динамике численности народонаселения, дает анализ возрастной структуры популяции. В развитых странах благодаря высокому уровню здравоохранения значительно увеличилась продолжительность жизни, что сказалось на возрастном составе населения. В популяциях возросла доля пожилых людей, тогда как детей и подростков стало очень мало. Возрастная «пирамида» выглядит вовсе не как пирамида, поскольку основание ее очень маленькое. Принципиально другая ситуация складывается в развивающихся странах, где доля младших возрастных групп существенно больше, чем доля старших. Обеспокоенность вызывает положение в странах Африки, расположенных к югу от Сахары, особенно в Нигерии. Сейчас в ней проживает 158 млн человек, но к 2050 году ожидается уже 326 миллионов. Среднее значение коэффициента фертильности — 5,7, а на долю тех, кому еще не исполнилось 15 лет, приходится 43 % численности популяции. Сложившаяся ситуация может измениться только тогда, когда начнет снижаться рождаемость.
Воздействие ранних цивилизаций на экосистемы Земли: избирательный характер уничтожения животных, пирогенное влияние, сведение лесов¶
Наиболее часто высказывается мнение о том, что человек с древних времен, когда он был еще собирателем и охотником, находился в более или менее органическом единстве с окружающей природой, почти не влиял на ее жизнь и полностью зависел от нее. В последующем он все более выходил из-под этой власти, постоянно нарушая былую гармонию общества и природы.
Действительно, интенсивность преобразований природы человеком последовательно возрастала. Наряду с этим сокращалась длительность этапов общественного и технического развития. Так, древний каменный век — ранний палеолит — длился сотни тысячелетий, поздний палеолит — несколько десятков тысячелетий, неолит — тысячелетия. Однако не следует забывать, что даже небольшие по силе воздействия, продолжающиеся длительное время, могут повлечь очень существенные последствия.
Один из самых ранних хозяйственных типов в истории человечества складывается из охоты на медведя и крупных копытных (лось, дикая лошадь, благородный олень, косуля, тур и другие), рыбной ловли, добычи водоплавающей птицы, сбора насекомых, съедобных моллюсков, растительной пищи (ягод, плодов, семян, желудей, корневищ, лесных и водяных орехов и так далее). Группы охотников-собирателей вели полуоседлый образ жизни. Ранненеолитовые поселения, как и позднемезолитовые, имеют культурные слои малых мощностей и размеров (50–200 м2), свидетельствующие о недолговременном существовании и о немногочисленности их обитателей, ведущих подвижный образ жизни. Анализ археологических материалов показывает, что численность групп охотников-собирателей достигала 20–25 человек. Каждая группа занимала обширную площадь, размеры которой зависели в первую очередь от наличия объектов охоты и съедобных растений. Средний показатель плотности населения для охотников-собирателей принимают за 0,05 чел./км2. Общая численность населения Земли в мезолите (10 тыс. лет назад) была около 3,5 млн человек, а к концу мезолита (6 тыс. лет назад) достигала, по мнению разных ученых, от 10 до 26,5 млн человек.
По некоторым данным, первобытный охотник тратил на добывание пищи в среднем 2–4 часа в день. О духовной жизни и относительном материальном благополучии его свидетельствуют великолепные наскальные рисунки, оставленные в разных районах мира. Районы с теплым и влажным климатом, богатые жизнью характеризовались большей плотностью обитания человека. На стоянках древних людей обнаружены каменные орудия труда, использующиеся для выполнения 30–40 функций, и кости животных. Проявлялась узкая специализация племен, охотившихся почти исключительно или на мамонтов, или на северных оленей, или на диких слонов, или на бизонов.
Универсальное оружие охоты — огонь использовался уже сто и более тысячелетий назад, что имело далеко идущие экологические последствия. Вполне приемлемой представляется гипотеза о том, что за вымирание некоторых видов фауны раннего плейстоцена, происходящее в результате разрушения среды обитания и источников питания, отвечает бесконтрольное использование огня, а не истребление человеком этих животных при помощи ручного оружия.
Первобытные общества охотников-собирателей соответствовали экосистемам, в которых они жили и являлись их частью наподобие других всеядных животных. Главное экологическое различие между людьми и другими видами животных состояло в использовании огня. Этот источник энергии дополнял энергию, потребляемую через пищу и необходимую для поддержания обмена веществ.
Рассчитали, что для существования одного человека в среднем палеолите требовалось около 2,1 × 104 кДж (5 тыс. ккал) в сутки. Они складывались из 1,3 × 104 кДж (3 тыс. ккал), получаемых с питанием, и 0,8 × 104 кДж, что давал очаг, с помощью которого готовилась пища и обогревалось жилье.
В нижнем палеолите, до того как человек стал использовать огонь и когда численность населения Земли не превышала 10–20 тыс. человек, годовое суммарное потребление энергии находилось в пределах (4,6 × 107)–(8,8 × 104) кДж.
В начале среднего палеолита численность жителей планеты достигла 200–300 тыс. и суммарное количество потребляемой всеми людьми энергии составляло (1,6–2,3) × 109 кДж/год.
В верхнем палеолите, когда на планете проживало около 1 млн человек, количество потребляемой энергии увеличилось до 9,2 × 109 кДж/год.
Следовательно, уже в верхнем палеолите использование огня в хозяйственных целях и рост численности жителей привели к увеличению энергопотребления на планете по сравнению с начальным этапом становления человечества в 100 раз. Подобный энергетический баланс первобытного общества не мог нанести существенного удара среде, хотя локальные нарушения природных комплексов могли быть достаточно заметными. Использование огня приводило к лесным и степным пожарам. Отходы жизнедеятельности охотников-собирателей быстро утилизировались природой, так как они, во-первых, были невелики по объему, во-вторых, распределялись по большим территориям из-за кочевого или полукочевого образа жизни. Ориентировочные расчеты показывают, что общее годовое количество нечистот от группы в 20 человек, равное примерно 10 т, рассеивалось по территории площадью около 400 км2. Прочие отходы охотников составляли органические остатки пищи и одежды из шкур, а также каменные сколы, образующиеся при изготовлении каменных орудий. Иными словами, охотниками — собирателями в природную среду не привносилось ничего для нее чужеродного. Продолжительность их жизни составляла в среднем 26 лет.
Более достоверно можно судить о взаимодействии с природой людей среднего палеолита — неандертальцев.
Неандертальцы обладали большой физической силой, их небольшие группы могли справляться с самыми крупными и свирепыми животными. Появление ледниковых покровов в северном полушарии сопровождалось серьёзным смещением природных зон, перестройкой экосистем. Неандертальцы были охотниками и собирателями. О собирательстве находок очень мало. Предположения об экологическом давлении приходится делать очень осторожно. Возможность такого давления определяется очень тонкой регуляцией природных экосистем. Происходит сопряжённое изменение численности хищников и жертв, представителей разных трофических уровней питания. Появление таких крупных и умелых хищников, как неандерталец, должно было сказаться на численности и распространении целого ряда крупных млекопитающих. «Выедая один пищевой пласт», человек переключался на другой. Влияние человека — прямое (преимущественно охота) и косвенное (огонь) — сказывалось прежде всего в крупных речных долинах, на морских и озёрных побережьях. Более или менее нетронутыми оставались Америка, Австралия, крупные и мелкие незаселённые острова.
Неандертальцу суждено было пережить мощную волну похолоданий, первую фазу валдайского оледенения. Затем последовало общее потепление климата — в промежутке между двумя стадиями похолодания. Неандертальцы сравнительно быстро исчезли, на смену пришли люди современного вида — кроманьонцы (ранние сапиенсы). Мягкий климат благоприятствовал их расселению. Эпоху кроманьонских охотников, собирателей, рыболовов, ещё не знавших металлов, называют поздним палеолитом (около 40 тысячелетий назад). Его формирование шло быстро — завершилось в течение нескольких тысячелетий, никаких других всепланетных природных явлений в этот период не отмечено.
В эпоху кроманьонцев произошло небывалое в природе явление — распространение одного вида живых организмов практически на всей обитаемой суше, во всех зонах. Кроманьонцы заселили Америку, Австралию, проникли в Субарктику. Ни один вид не имел такого распространения, не говоря уж о способности влиять на несколько трофических уровней экосистем (хищные и травоядные животные).
Расселению человека способствовало оледенение: вода на суше в виде ледниковых покровов, уровень Мирового океана понизился на десятки метров. Британские, Японские, Зондские и другие острова были частью континентов. Северо-восточная Азия и Северо-западная Америка соединялись широким перешейком. Кроманьонцы по суше перешли в Америку. Для освоения Австралии и Океании они овладели навигацией. Для некоторых видов животных, некоторых экосистем это оказалось весьма существенным фактором. Наступает новый этап развития человечества и биосферы. Люди употребляют новые виды рыб и птиц в невиданной прежде степени. В мир пришла техника (выжигание угля, обжиг гончарных изделий, обработка каменных орудий, оружия, приспособлений из костей, рогов животных и так далее)
Оценить масштабы влияния кроманьонцев на природу косвенно можно на территориях, заселенных лишь в позднем палеолите (Америка, Австралия, острова).
За последние десятки миллионов лет в Новом Свете произошло, пожалуй, только одно крупное и катастрофически быстрое вымирание млекопитающих. Заключительная фаза последнего оледенения сопровождалась поначалу обогащением видового состава фауны. Несмотря на морозный климат и гигантские ледники, в Америку из Евразии перекочевали несколько родов крупных млекопитающих (овцебык, сайга и другие). Общее потепление климата и таяние ледников благоприятствовали повышению продуктивности экосистем. Численность животных должна была возрасти. Но случилось нечто противоположное. За период 2 тыс. лет исчезает несколько десятков родов (30 видов) животных, вскоре после прихода сюда человека. Влияние человека не коснулось растений, беспозвоночных, морских позвоночных. Большинство видов исчезло внезапно между 11 тыс. и 8 тыс. лет, вымирание мамонта в районе Скалистых гор и Высоких равнин произошло примерно 11 тыс. лет назад. Анализ пыльцы и спор растений показывает, что 10 тыс. лет назад растительность была сходна с современной. Никаких климатических и геологических катастроф не было. Исчезли верблюды, лошади, ленивцы, мускусные быки, винторогие антилопы, отдельные виды кошек, оленей, два вида мамонтов — все это результат активной охоты первобытного человека. Увеличение размера крупных животных, достигнутое в ходе эволюции, снизило в той или иной степени риск нападений хищников. С исключением этого важного фактора уменьшения численности вида сделало возможным и целесообразным заметное сокращение рождаемости. Когда человек верхнего палеолита начал охоту на этих животных, они стали вымирать вследствие низкой рождаемости. Животные Нового Света оказались беззащитными перед человеком, погибали от пожаров, из-за нарушения экологического равновесия (хищники при уничтожении травоядных). Исчезли либо самые «доступные» при охоте, либо самые ценные виды животных. Мелкие млекопитающие совсем не пострадали. Сохранились бизон и северный олень: подвижный бизон — за счет расширения человеком степей и лесостепей (огонь уменьшил площадь лесов), оленя хранил суровый климат. Примерно в то же время во многих других крупных регионах исчез целый ряд крупных млекопитающих ледникового периода, до этого достаточно многочисленных. Благоприятные условия для охоты по сравнению со странами Европы и Азии, где дикие животные соприкасались с человеком на протяжении сотен тысяч лет и выработали различные способы защиты от преследований, определили быстрый прирост и заселение человеком всей Америки.
В научных трудах была обоснована и другая модель перестройки ландшафтных комплексов суши на рубеже плейстоцена и голоцена. На протяжении всего плейстоцена во всех климатических поясах растительность никогда не имела столь резко выраженной зональной структуры, как в голоцене. Границы растительных зон были гораздо более размытыми, леса имели парковый облик и так далее. Во всех климатических зонах мозаичность фитоценозов была, по-видимому, выше, чем в голоцене. Некомпенсированному вымиранию крупных животных — фитофагов на рубеже плейстоцена и голоцена предшествовал период активного совершенствования охотничьих орудий. Увеличение численности палеолитового населения, совершенствование орудий и техники специализированной охоты на крупных травоядных животных привели к тому, что численность их популяций стала снижаться на всех материках 15–20 тыс. лет назад. К рубежу плейстоцена и голоцена численность «ключевых фитофагов», в том числе мамонтов, шерстистых носорогов, гигантских ленивцев Южной Америки и других, катастрофически снизилась. На большей части территории континентов они исчезли, а на остальной уже не могли исполнять роль эдификаторов растительных сообществ из-за низкой численности. В отсутствие «ключевых фитофагов» стали формироваться ценозы намного менее мозаичные. Возникла хорошо знакомая нам система ландшафтных фитоценозов с четко выраженной зональной структурой. Это привело к глубокой перестройке фаунистических комплексов, для многих представителей которых не оказалось места в новой структуре биоценозов. Часть животных, наиболее тесно связанных с мозаичностью ранее существующих биоценозов вымерли, другие сохранились лишь в той части своих прежних ареалов, где условия существования остались достаточно приемлемыми для них.
Особенно глубокие изменения претерпели фаунистические комплексы тех зон, где этот процесс обострился из-за резких изменений климата. Единый плейстоценовый комплекс «мамонтовой фауны» Евразии фактически перестал существовать. Отдельные его представители сохранились в разных климатических зонах — от арктических пустынь до пустынь Средней Азии и Монголии, но в настоящее время области их распространения не только не перекрываются, но и даже не контактируют друг с другом.
Гибель основных видов охотничьих животных и коренное изменение облика ландшафтов потребовали полной перестройки хозяйственного уклада. В мезолите основой существования стали относительно малоспециализированная охота практически на все виды доступных животных, рыболовство, собирательство.
Другую причину гибели крупных млекопитающих видят в резком контрастном изменении климата. Быстрая деградация вечной мерзлоты, смена экологических условий — и животные не смогли приспособиться. Однако даже в этом случае участие человека в гибели млекопитающих не вызывает сомнений.
Влияние человека каменного века можно наглядно проследить на изменении природы Тасмании и Австралии. По археологическим находкам территория острова Тасмания заселена около 30 тыс. лет назад при опускании уровня океана в эпоху оледенения. Тасманцы занимались охотой, рыболовством, собирательством. С помощью огня они перестраивали свои ландшафты, с целью увеличения площади открытых пространств и диких пастбищ. Влажные леса уступили место кустарникам, саванам, степной и болотной растительности. Рост населения был ограничен экологической зависимостью в системе «хищник-жертва», постепенно появились признаки упадка материальной культуры.
В Австралии охотничьи племена постоянно кочевали. Каждая группа ежегодно выжигала около 100 км2 лесов, саванн, степей — целенаправленно или невольно. Тысячи таких групп могли за 30 тысячелетий десятки раз выжечь растительность всего континента. Создавались огромные пирогенные территории, что в долговременной перспективе вело к обеднению биоресурсов, опустыниванию. Процесс обострялся в периоды засух. Леса — хорошие стабилизаторы почвенной влаги, их уничтожение вызывало большую потерю воды, иссушение и эрозию почв. Разрежение растительного покрова сначала благоприятствовало увеличению численности крупных животных. Однако через века и тысячелетия, когда земли эродировали, для людей наступили трудные времена. В истории аборигенов после подъема материальной культуры наметился явный регресс, приблизительно тогда, когда были уничтожены крупные сумчатые континента.
Считают, что окончание культуры палеолита в Европе, возможно, было, в известной степени, результатом неразрешимого противоречия между созданной человеком верхнего палеолита техникой массовой охоты на крупных животных, обеспечившей временное изобилие пищи и сделавшей возможным увеличение численности населения, и ограниченностью природных ресурсов для этой охоты, которые через некоторый период времени оказались исчерпанными. Причины этих экологических нарушений заключаются в необычайно высокой скорости эволюции человека по сравнению со скоростью эволюции преследуемых им животных.
Переход к мезолиту имел болезненный характер для первобытного общества и сопровождался временным сокращением численности населения.
Средиземноморье было очагом формирования кроманьонцев и длительного хозяйствования первобытных охотников и собирателей. 10–20 тысячелетий назад Средиземноморье было покрыто почти сплошными густыми лесами. В пору мезолита началось активное уничтожение лесов — не столько вырубками, сколько преднамеренными и непреднамеренными пожарами. Расчленённый рельеф территории способствовал эрозии почв, появляются деградированные ландшафты. Нарушается водный режим территории: разливы рек и наводнения после дождей сменяются безводием сухих русел и недостатком воды в засухи.
В Африке на территории пустынь, полупустынь и сухих степей когда-то жили люди. Остатки животных и растений, русла водоемов свидетельствуют о том, что земли не испытывали острого дефицита воды. Считается, что под действием ледников средиземноморская растительность была оттеснена на юг — в Сахару. Озеро Чад занимало площадь в 8 раз больше современной. Со времени кроманьонцев в Африке не было экологических ниш, не занятых человеком (кроме, может быть, глубинных районов влажных лесов). Учитывая активность человека, вряд ли можно сомневаться в серьезных изменениях экосистем. Наибольший урон приносили пожары. Разрушился почвенный покров, понизился уровень грунтовых вод, пришли в движение пески. Пирогенное воздействие, конечно, вызывало необратимые изменения неустойчивых лесных экосистем в условиях субаридных климатов.
Колебания численности хищников и жертв — известная экологическая модель. В этом отношении человек оказался особым компонентом экосистем. Уничтожая крупных млекопитающих, он избегал вымирания, ему не нужно было изменяться биологически, приспосабливаясь к новому «пищевому пласту», достаточно было выработать соответствующую технологию. Он переключался на добычу мелких и юрких зверей, птиц, рыб, собирал растения. Однако постепенно создаются предпосылки для кризиса охотничьего хозяйства, присваивающего ресурсы без возобновления.
Антропогенная эволюция экосистем
Древние очаги земледелия приурочены к современной засушливой области обедненных, опустыненных ландшафтов. Первоначальными районами освоения были предгорья и низкогорья. Здесь благоприятные для земледелия участки сравнительно невелики. Сведение леса человеком разрушало почвенный покров, резко усиливало сток поверхностных вод, возникали катастрофические паводки, отрицательно влияющие на растительность и животный мир речных долин и озерных впадин и на жизнь обитающих здесь племен земледельцев. Уменьшение грунтового стока обедняло и природные экосистемы. Постепенно изменялся и микроклимат в сторону усиления контрастности температур и степени увлажненности. Измененные и деградированные территории увеличивались по площади. Все это происходило на фоне обычных климатических колебаний, не обязательно направленных на опустынивание.
В средиземноморье активные формы современного карста, как правило, вторичны. Они развиваются на более крупных формах древнего карстового рельефа. Этот древний рельеф был переработан в виллафранке (самом конце плейстоцена) при более влажном климате. Позднее процессы карстообразования постепенно затухали, происходило «одряхление» карста. Многие большие полья были заполнены аллювиальными отложениями. Здесь выросли леса и сформировалась поверхностная гидросеть. В течение тысячелетий человек сводил леса. Перевыпас и земледелие привели к деградации почвенного покрова, который удерживал атмосферную влагу. Обнажились коренные из-вестняки. Водотоки ушли под землю, в лабиринты древней карстовой гидрографии. На больших площадях образовались «самые карстовые» ландшафты на земном шаре — известковые пустыни.
В эпоху неолита (средний каменный век) человек впервые выступил как активная геологическая сила. Это выразилось в интенсивности и разнообразии форм воздействия на среду: скотоводство, земледелие, строительство крупных поселений, активное уничтожение лесных массивов, строительство ирригационных систем, шахт. Повышение благосостояния вызвало рост численности населения планеты до нескольких десятков миллионов человек, что еще усилило масштабы воздействия на природу. Как творец «второй природы» человек оказывается в центре мироздания. Создание искусственных ландшафтов, разрешение противоречий с природой вели к появлению математики, письменности, к выделению различных социальных групп. Чем совершеннее становились окультуренные растения, животные, экосистемы, тем больше отчуждались от естественной среды, требуя для своей устойчивости и совершенствования знаний, затрат энергии.
Генеральное наступление земледелия из эродированных предгорных районов шло в направлении низменностей, долин крупных рек. Первоначально были освоены Месопотамия, Двуречье, затем долины Нила, Инда, бассейн Хуанхэ, междуречье Амударьи и Сырдарьи.
Сначала потребность растущего населения удовлетворялась освоением целинных территорий. Позднее появились ирригационные системы, системы защиты от наводнений. Для расширения территорий полей и пастбищ успешно применялся огонь, уничтоживший леса в полуаридных условиях. Сведение леса в верховьях рек делает паводки более контрастными, проходят чаще селевые потоки, разрушаются почвы, заиливаются русла — усиливается опасность наводнений.
Внедрение индустрии меди сказалось на состоянии окружающей среды, так как для выплавки использовалась древесина, что ускорило деградацию лесных массивов.
В отношении численности населения на заре аграрной культуры оценки демографов расходятся. По разным расчетам по сравнению с неолитом численность возросла в несколько раз (ко 2-му тысячелетию до н.э. — 50–133 млн человек). С середины 4-го тысячелетия до н.э. на Ближнем Востоке и в Египте города и поселения стали объединяться и формировать государства, которые превращались в рабовладельческие империи. При изучении проблемы численности населения ученые пришли к выводу, что в эпоху аграрной культуры колебания в численности людей тесно связаны с изменением количества пищи и что на протяжении истории землевладельческих обществ наблюдались резкие пики смертности — до 150–300 и даже 500 на 1000 человек. Иногда они приходились на периоды войн, но чаще были результатом голода и эпидемий.
Энергопотребление на душу населения в античном мире лишь немного превысило уровень эпохи неолита, увеличившись с 4,6 × 104 до 5 × 104 кДж/cут. Казалось бы, что такой небольшой прирост энергопотребления не в состоянии значительно изменить общественное производство и обеспечить энергетический базис роста численности населения. На самом же деле античные науки и культура достигли небывалого расцвета. Крайне низкий уровень потребления энергии был у рабов. Они сами стали поставщиками мускульной энергии наряду с домашними животными. Примитивное производство концентрировалось в отдельных локусах, там и росла удельная плотность энергопотребления. На этих участках удельное энергопотребление достигало 2,1 × 1010 кДж/кв2 в год.
Литература¶
-
File:BENNU’S JOURNEY - Early Earth.jpg. (2024, July 17). Wikimedia Commons. Retrieved 16:11, August 8, 2024 from https://commons.wikimedia.org/w/index.php?title=File:BENNU%E2%80%99S_JOURNEY_-_Early_Earth.jpg&oldid=899962140. (NASA's Goddard Space Flight Center Conceptual Image Lab) ↩
-
File:Hadean.png. (2024, March 22). Wikimedia Commons. Retrieved 16:14, August 8, 2024 from https://commons.wikimedia.org/w/index.php?title=File:Hadean.png&oldid=862671536. (Художественное представление Гадейского эона. АвторЖ Tim Bertelink 🡥) ↩
-
https://stratigraphy.org/ 🡥. Страница с интерактивной схемой, схема в формате .xlsx и другие — https://stratigraphy.org/chart#latest-version 🡥. ↩